Würfelkunde Ende: Haare Schick und Big Bang putzen mir den BiXnom blank
In der Abteilung Ulk geht es um Unnützes, aber wegen des Bezugs aufs Wasser, musste ehrgeizig gesucht werden - überspringen! Die errechnete Streckenteilung wurde auf das runde Spielfeld direkt angewendet. Man erhält einen Kern für das Potential im Radius 1 und eine Kontakthülle der Breite = +0,2533. Dieser Kreisring ist als BiXnom-Wassermodell zu verstehen. Je nach Datenquelle zum Wassermolekül ist zu erwarten, dass das Wasserstoffatom mit seinem Radius kontaktsuchend aus der Kontakthülle herausragt. Aktuell ist bei Wikipedia als Bindungslänge (Mittenabstand der Atome O und H) = 95,84 pm zu finden. Die Größe des Wasserstoffatoms im Molekül ist in dieser Quelle z.Z. noch nicht vermerkt.
Bild Siebert: Wassermodell
Das Kreisringmodell kann einzeln oder auch das Kugelschicht sinnvoll genutzt werden, beispielsweise auch für Baustoffe, die mit Wasser aufgemixt wurden. Eine hinterlegte Bienenwabenstruktur sorgt dafür, dass man für die dimensionslose Betrachtung eine Masse einrechnen kann. In den Kreisringen lagert beispielsweise Wasser und mittig im Kern der Zement umringt von Zuschlagstoffen. Ein Mikro-Betonmischer kann dann nach der Rezeptur Frischbeton erzeugen. Ohne weitere Besonderheiten kommt bereits die sichere Heimwerkermischung 1:3 heraus. Die Bienenwaben könnten auch noch ein Statik-Modell beinhalten, so dass dann Bauwerke aus dem Drucker gemixt würden.
Die Kontakthüllenbreite lässt sich in einen Sinus von einem Kontaktwinkel Phi umrechnen: sin Phi = 0,253314 bzw. Phi=14,6737°. Der Kontaktwinkel soll nicht zwingend etwas mit Reibung zu tun haben. Im Wasser entsteht erhebliche Reibung, wenn hochturbulente Wasserballen zusammenstoßen oder Wasser sich auf krummen Strombahnen aus Öffnungen quält. Es kommt nun kein Kontaktwinkel von nahezu Null heraus, sondern ein Wert, den man sonst als Reibungswinkel für Tonmineral kennt. Aus der Mikrostruktur kann man es sich aber erklären: Damit ein hexagonales Prisma holprig ewig rollt, muss es sich auf einer schiefen Ebene mit einer Neigung von 6,58° befinden [29. Internat. Physikolympiade 1998 in Reykjavik/Island; Aufgabenbeschreibung Prof. H. Mayr, Universität Wien , Plus Lucis S1/99, Seite 26]. Wassermoleküle schließen sich zu hexagonalen Gruppen (Clustern) zusammen. Setzt man in dieses Gerüst das Wassermodell ein, dann ist eine größere Hubhöhe erforderlich, die eine größere Rampenneigung erfordert. Rampenneigung ist wie eine Böschungsneigung und diese hat unter Wasser eine Ähnlichkeit mit dem Reibungswinkel.
Bild Siebert: Bindungswinkel
Die Vorstellung ist somit, dass Wasser imaginär ständig auf einem Energiegefälle abrollt, sofern es sich nicht um Eis handelt. Das Wasser hat diese Rampe schon eingebaut. Das Wassermodell rollt weicher als ein harter Klotz bis auf sein Kern ab und hier ist dann ein Abrollwinkel von ca. 6,5° vorhanden. Im Hexagonalen Raster sind es 6,655..° und als Kreistangente 6,487...° (i.M. 6,57°). Das Wassermolekül steht hierbei in einem Tetraeder in einem dreieckigen Grundriss. In einer anderen Ansicht wird das Tetraeder sichtbar. Wasser weicht mit 104,45° vom Tetraederwinkel=2*arctan (Wurzel(2)) = 109,47° ab. Außer dem Unterschied von 2 freien und 2 gebundenen Bindungen, geht der Tetraederwinkel nur auf ein Quadrat und nicht auf einen Kreis zurück, er rollt mit einer Anpassung besser. Auch in dieser Koordinatenachse ist ein Zusammenhang mit dem Abrollen festzustellen.
Für andere Stoffe verändert sich der Kreisring, sofern es überhaupt ein Ruhemodell dafür gibt. Es interessiert die Ringbreite für Eis und für Granit nur der Reibungswinkel selbst. Dazu wird der Radius des Spielfeldes in der rücksichtslosen Variante einfach durch den Formfaktor f = Stoffreindichte/Wasserdichte geteilt. Damit das Binom hier noch mitspielen kann, muss sich die Breite b im Binom verändern. Stoffe können sich durch Kristallisation anpassen. Dichten < 0,6 sind im Modell nicht möglich. Der Kontaktwinkel ist bei einer Dichte von 1,2533.. gleich Null. Richtung Wasser dann positiv und Richtung Granit dann negativ. Kontaktwinkel größer 45° sind nicht möglich, weil man sich nicht an den eigenen Haaren aus dem Sumpf ziehen kann. Die im Wassermodell unbeachtete 2. Erscheinungsform (TYP2 ), ergibt Abstandsverhältnisse, die für eine Sinusfunktion bzw. einen Winkelergebnis unbrauchbar sind. Die Zunahme der absoluten Ringdicke in beide Richtungen (größeres Eisvolumen / Granit mehr Festigkeit) ist verständlich. Für die Biologie muss man sich bei gleichem Ergebnis eine einfühlsame Vorstellung machen, denn die Meduse (Qualle) wiegt im Wasser nichts. Hierzu wird der Radius des runden Spielfeldes nicht durch den Formfaktor geteilt, sondern 1. wird die runde Fläche (der Medusenschirm ist rund) durch den Formfaktor geteilt und die Rechteckfläche des Binoms gleichzeitig mit dem Formfaktor multipliziert. Diese Multiplikation wirkt als 3. Dimension. Zieht sich der Schirm zur Bewegung reibungsmindernd zusammen, klappt er nach hinten becherförmig weg.
Bild Siebert: Formfaktor(Reindichte) und Kontaktwinkel
Wenn etwas nur gepresst wurde wie Graphit zu Diamant, könnten sich Stoffe mehr oder weniger zufällig übereinander in einer um jeweils Wurzel(2) versetzen Kurve finden lassen. In einen Würfel passen 3 Tetraeder hinein, so dass noch 2 Kurven möglich sind. Es geht also nicht über 45° nach rechts hinaus, dafür aber höher (Minusbereiche als Absolut ins Positive spiegeln). Wenn das noch nicht reicht, geht es mit künstlichen Gelenken und Smartphone für Körper und Geist immer noch weiter nach oben. Für die üblichen rolligen Erdstoffe (z.B. Granit mit 2,65 g/cm³) ergeben sich bekannte Reibungswinkel von 31-32°. Werden unterschiedliche Porenvolumen der Erdschichten noch berücksichtigt, entsteht eine Berechnungsformel für den Reibungswinkel. Dieser wird z.B. für die Berechnung des Erddrucks auf die Kelleraußenwand benötigt. Das Gegenteil einer Kugelpackung ist die flockige Struktur ohne Kugeln, die für die bindigen Stoffe (Ton) geeignet ist.
Bild Siebert: Die Erdform
Bei einem Reibungswinkel von 45° hat die vertikale Druckkraft maximal eine gleichgroße horizontale Reibungskraft erzeugt – mehr geht nicht. Die BiXnom©-Stoffgrenze für 45° liegt bei 4,2791 g/cm³. Die mittlere Dichte der Erde liegt bei 5,515 g/cm³, so dass sie nach innen geschichtet sein muss. Mit der Idee die Stoffe einfach enger in das selbe Volumen zu stopfen, kommt man nicht an die Metalle heran, dann hilft der Wechsel von der kleinen Form zur großen Form „Erde“. Darin zirkulieren die geschmolzenen Metalle wie Wasser. Wieviele Schichtungen hat die Erde? Es sind 8, wobei es 7 Ringe sind und der 8. ist der Erdmittelpunkt. Woher wissen wird das? Die Hüllendicke in der Bixnom-Kurve ist bei 1,2533 g/cm³ gleich Null. Hier liegt der blanke Kern des Wassermodells. Dieses ist somit die Stapelhöhe vieler BiXnom©- Kurvenkopien. Am rechten Ende jeder Kurve liegt dann immer der 45°-Grenzwert: Erreichbare Dichte für den Erdmittelpunkt (7. Stapelung) = 4,2791 g/cm + (7*1,253314) = 13,05 g/cm³. Die Dichte im Erdmittelpunkt beträgt tatsächlich 13,00 g/cm³. Der Schichteninhalt ist nun bekannt, aber wie findet man das Ringmodell für Erde? Man kann nur ganz einfach mit dem Wassermodell anfangen. Nun stapelt man an der äußeren Hüllentangente, 8 Hüllenschichten auf. Es ergeben sich Dreiecke, in denen die wachsenden Hypotenusen die Radien der geschichteten Ringe sind. Nun fragt man sich, was diese kleine Lücke aus dem Unterschied von Sinus und Tangens am Hüllenrand bedeuten soll. Kann die Lücke weg? Ja, natürlich und das ganze Wassermodell selbst kann auch verschwinden? Weshalb? Das konstruierte Ringsystem wird so gespiegelt, dass der äußere Ring jetzt zum Kernmittelpunkt wird. Die Erde ist das Gravitationszentrum, das Wassermodell wird nicht mehr benötigt. Das Wasser liegt bei dieser Betrachtung kernlos, und zwar jetzt im Modell außen, in der Erdkruste als Ozeane herum. Das Ringsystem wird durch einen Maßstabsfaktor auf den mittleren Erdradius von 6371km skaliert. Irgendwelche Ringe kann jeder zeichnen. Erfolg ist natürlich nur dadurch beschieden, dass einzelne Ringe auf wissenschaftlich bekannte Grenztiefen treffen. Wenn nicht jeder Ring auf eine bekannte Grenze trifft, liegt das daran, dass die Erde nur eine begrenzte Anzahl von Stoffen enthält, und die fehlenden Stoffe durch Zirkulation der Schmelzen simuliert werden muss. Erst der innere Kern ist fest. Die BiXnom©-Erde kann präzise gezeichnet werden. Alle gestapelten Schubladen des Schrankes „Erde“ zusammengezählt, würden die Erdmasse als zu schwer ausweisen, weil eine neue Schublade leichter beginnt als die zuletzt voll gefüllte. Die noch zu suchende Formel müsste eine stetige Massensteigerung enthalten.
Bild Siebert: Die Bixnom-Erde
Der BiXnom-Meter
Don't Be Cruel - Billy Swan
Ein Meter ist die Strecke, die das Licht im Vakuum während der Dauer von 1/299 792 458 Sekunde zurücklegt. Die Strecke entspricht seit 1799 dem Urmeter. Dieser Platinstab entsprach nach den damaligen Messungen dem zehnmillionsten Teil der Entfernung vom Nordpol zum Äquator. Falls die Erde alle Massen vorbeiziehen ließ, die ihr mathematisch nicht genügten, lassen sich aus der Meter-Definition auch Natur-Informationen herausziehen:
Die Bogenlänge von 10000 km hat eigentlich keinen Naturbezug. Ein irgendwie auf einen Haufen geworfener Bindfaden gleicher länger hat auch keine Aussage. Es war also nur eine sowieso nötige Erdvermessungsaufgabe, denn welche ungefähre Länge als Meter gelten sollte, war der Wissenschaft anhand des mathematischen Pendels (Sekundenpendel) bereits bekannt. Ein Naturzusammenhang über eine Strecke ist aber durch den Wasserwürfel für die Kilodefinition gegeben, so dass hier im Kunstprojekt der Kreis erst aus einem Quadrat des gleichen Umfangs entwickelt wird. Beachte: „umfangsgleich“, ein Kreis schließt damit eine extrem größere Fläche ein. Bogenlänge Erdquadrant = Kantenlänge Einheitsquadrat = 10.000,000 km.
Für die Quadratfläche wird ein flächengleicher Ersatzkreis gefunden, mit dem sich im Quadrat durch Schnitt mit den 45°-Diagonalen eine 9-feldrige BiXnom-Teilung finden lässt. Radius Einheitsquadrat-Ersatzkreis RE= Wurzel [(10000^2)/Pi] = 5.641,895835 km < fiktiver Erdradius 6.366,197724 km. Vom Zentrum zum Mittelpunkt eines Eckquadrats M entsteht ein Radius R_ lokal= 6356,4818236715 (Kontakt zum Berechenbaren) und zum Endpunkt der Diagonale E der Radius R_global = 6396,5220478937 (Kontakt zum Unberechenbaren). Der Erdradius wird damit beidseitig umgrenzt. Der Erdradius ist mittig zwischen beiden Punkten zu erwarten, was aber knapp verfehlt wird. Der lokale Erdradius ist 61,04680015 km kürzer als der Erdradius und würde als Meterdefinition L = (10000-61,04680015)/10000= 0,99389532 m bedeuten. Diese Strecke sei nun die Stablänge des Sekundenpendels, was eine Erdbeschleunigung g = L / (T1/2 / Pi)^2 von g = 9,80935362 m/s² ergibt. Ein passender Ort dafür ist Paris, wo das Urmeter allerdings mit 1 m und leicht schrumpfend aufbewahrt wird.

Der größere globale Kreis würde eine etwas zu kleine mittlere Erddichte liefern. Es wird noch ein Dichte-Zuschlag benötigt. Die mittlere Erddichte wird ideell an einer volumengleichen Kugel mit der gleichen Anziehung ermittelt. Enthält die Formel den hier betrachteten Erdumfang U, ergibt sich als mittlere Erddichte = 1,5* g /(G*U). Mit Erdbeschleunigung g = 9,81 m/s², Gravitationskonstante G = 6,6732*10^-11 m³/kg*s² und U = Erdumfang 40.000 km ergeben sich 5,51 g/cm³. Für die abgeplattete Erde sind es ca. 5,517 g/cm³. Die mittlere Erddichte soll alternativ aus dem 1 Kilo „Wasserwürfel“ ähnlich gefunden werden: a) Für die globale Dichte wird der irrsinnige Kontaktwinkel von +14,67371281° für 1 g/cm³ angesetzt. Märchenhaft gekontert: Die Erde kann ihr Wasser nur mit einem entgegengesetzten Negativwinkel als Mineralbereich halten. b) Für den lokalen Dichtezuschlag soll der vorstellbare Abrollwinkel von ca. 7° gefunden werden, wie er für ein hexagonales Cluster von Wassermolekülen zu erwarten ist. Idee: Die Erde zieht als Flut zurück, was der Mond an Ebbe bewirkt hat. Die BiXnom-Formel reicht für die Erdkruste nur bis 4,2791g/cm³, so dass bis in die 4. BiXnom-Schicht gestapelt werden muss. Der Konterwinkel +14,67371281° ergibt aus der BiXnom-Formel eine Dichte von 1,6785g/cm³. Jetzt wird der Basiswert von 3 Stapelschichten darunter geschoben. Globale Dichte =1,6785g/cm³ + 3* Wurzel (Pi/2) g/cm³= 5,4384 g/cm³. Ein Drehwaageversuch von 1838 durch F. Reich ergab 5,44 g/cm³. Dichte-Zuschlag = 1 g/cm³ * Formfaktor * Volumenfaktor: Die Hüllendicke wird auf den Ersatzkreis für das Erdquadrat bezogen, um ein BiXnom-Modell mit Einheitsradius und Hülle zu erhalten. Mit der Formel „Formfaktor f = [Wurzel (Pi/2)] /[[sin (Kontaktwinkel)] +1]“ wird der Formfaktor und dann die Differenz für beide Kreise ermittelt zu: Delta = 0,00696339. Der Formfaktor ist dimensionslos, hat hier aber als g/cm³ erst eine Verwendung, wenn die Quadratfläche in Würfelform durch Faktor 10 und die Pendellänge in Abweichung zum Meter in Kubik angerechnet werden.
Dichtezuschlag = Delta * [10/(0,99389532^3)] = 0,00696339* ( 10/0,9817975) = 0,070924908 g/cm³
BiXnom-Dichte = Globaldichte plus Lokalzuschlag = 5,4384 + 0,070925 = 5,50932 g/cm < 5,51 g/cm³
Genau das musste natürlich herauskommen – Wasser bleibt liegen, ist aber auch ein knappes Gut!
Tauchtiefe = Meeresspiegel bis lokaler Hüllkreis = 6366197,7 - 6356481,8 = 9.716,0 m tief. Zufällig passend: Atlantik = 8486 m, Pazifik = 10.994 m und beide gemittelt = 9.740,0 m. Umgekehrt stünde über dem Meeresspiegel ein gewaltiger „Wasser-Berg“, der mit der max. Krustendichte gleichwertig mittels 7000er Gebirgsketten gebildet wird: 30.302,4323 m * (1,0 g/cm³ / 4,2791g/cm³) = 7081 m hoch. Alles nur aus 1 Meter!
Als wesentlich wurde gefunden, dass die 4 Stapelebenen zu 4 Quadranten passen. Im BiXnom-Vakuummodell liegt dort jeweils 1 BiXnom-Basismodell und für die Biologie auf der Erdoberfläche wird nur 1 Basismodell betrachtet. In beiden Betrachtungen fallen Maße direkt als Naturmaße an. Das verwundert, muss aber kein Wunder sein [dat. 12.08.2020].
Staple keine Biolog*innen, ansonsten eine je Erdquadrant.
Der BiXnom-Masterpunkt E* ist durch das Sonnensystem natürlich realisiert und erklärt sich nur als Rückblick zum geometrischen Entstehungsgrund: Der Radius um den Global Point E „rX= 11,4152201192 km“ kann dimensionslos als BiXnom-Modell mit Kernradius 1 und Hülle = 0,14152201192 gelesen werden, was einen Kontaktwinkel arc sin (0,14115...) = 8,135928098° ergibt. Der Radius rX ergibt sich als Lösung einer quadratischen Gleichung. Der Master-Point E* konvertiert die Dauer des Erdumlaufs um die Sonne auf den Erdkreisumfang mit 360°. Anomalistisches Jahr (mittlere Bahnperiode der Erde) = 365,259 636 Tage. Anzahl Abschnitte = 365,259 636 /8,135928098= 44,8946489938 rd. 45 Stück. Etwas Zusatzzeit könnte noch durch das Bummeln am Erde-Mond-Schwerpunkt eingerechnet werden. Die 8 Masterpunkte bilden somit die Anzahl der Jahrestage auf einem Vollkreis ab: 8x45= 360°. Das Jahr hat 365,259 636 Tage* 24 Stunden/Tag* 60 Minuten/Std.* 60 Sekunden/Min. = 31.558.432,5504 Sekunden. Die Sekundenabschnitte werden bei Teilung durch Pi auf den Durchmesser geschaufelt, der sich dann ergibt zu 31558432,5504/ Pi= 10.045.361,0732567. Weshalb Pi? Eine Wissenslücke ist keine Kunstpause: Die Maßeinheiten vom Gravitationsmodell "Erde" werden für den Rahmen eines leeren Vakuummodells benötigt, um dessen Inhalt "Nichts" zu verstehen. Das Äquivalenzprinzip der Physik könnte die Situation evtl. erklären. Es sind also diese 10 Mio Einheiten, so dass 1 Einheit auf jeden Meter des Erdquadranten der Meterdefinition entfällt. Weg und Zeit stehen über die Geschwindigkeit im Verhältnis. Neuzeitlicher Vermessungswert zur Länge des Erdquadranten = 10.001,965 km. Die noch fehlenden 43 km "schmieren" wir nun ideel als Mond-Pudding auf die Erdoberfläche: Dicke der Mondpaste = Mondvolumen / Erdoberfläche = 220/5,1= 43,13 km. Wow! Leider kein Kunststück, weil keine Länge, sondern eine Dicke in Richtung des Radius. Somit aber als passender Bummeleffekt verwendbar. Wegen D = Umfang / Pi liegt die Zeitinformation auf dem Durchmesser. Weil das BiXnom-Vakuummodell aber aus 4 BiXnom-Basismodellen besteht, liegt die Zeitinformation auf dem Radius des Vakuummodells. Die Lichtgeschwindigkeit kann einfach wie ein Radius gezeichnet werden, was dann zur BiXnom-Lichtkugel führt (dat. 17.08.2020). Das Vakuummodell führt vom Erdkreis in den Mikrokosmos, wodurch die Genauigkeit extrem steigt. Das BiXnom-Wunder selbst erklärt: Drehen sich die Autoräder passend zur Kamerabildfolge, drehen sich die Räder des Autos scheinbar nicht, obwohl das Auto wie die Lichtgeschwindigkeit in km/h messbar ist. Weil die Lichtgeschwindigkeit dimensionslos durch 10^8 dividiert ist, verkleinert sich der Bildrahmen entsprechend. Für den Freundeskreis "Würfelkunde" sind dann Details im Bildrahmen noch 10fach kleiner als das Licht im Tunnel sichtbar - es ist die Wellenlänge des sichtbaren Lichtes von 380 -780 nm.
Der Goldene Schnitt der Erde und die Neigung der Erdachse - The Normal One
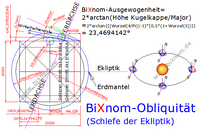
Die Neigung der Erdachse wird einer Kollision in der frühen Entstehungsphase der Erde zugeschrieben. Tatsächlich ist die Neigung trotz der Schwankungen in der Erdgeschichte von 21,8 - 24,4° über 40000 Jahre (Schönwiese 1995, S. 145; zit. in geo.fu-berlin.de) durchaus verständlich. Gemäß Triebkraft der Würfelkunde "möchte" die Erde nur das Würfelvolumen aus der Kantenlänge von 10000 km besitzen, muss aber gravitationsbedingt aufgrund der Kugelform einen ungeliebten Ballast bei einer bereits großen mittleren Erddichte mitschleppen. Die Neigung ist dann auch einfach wie eine Böschungsneigung zu sehen - was "abrutschte" ist halt weg. BiXnom-Neigungswinkel "Ausgewogenheit" = 23,4694142409°. Dieser Neigungswinkel ist praktisch mit 23,43661° auch vorhanden. Erdbeben, Gletscher-Eisschmelze, Grundwasserentnahmen, Kontinentaldrift etc. führen zu Schwankungen incl. Eiszeiten. Stärker geneigt, bringt im Sommer und Winter auf beiden Halbkugeln extreme Temperaturen. Weiterhin schwankt die Exzentrizität der Erdbahn um Sonne von kreisrund bis elliptisch alle 100000 Jahre. Die BiXnom-Ausgewogenheit ALPHA ist bei jeder Gravitationskugel universell gleich, weil der zu Grunde liegende Goldene Schnitt dimensionslos ist (dat. 24.08.2020). Der Frühling und die Schönheit sind im Universum nicht selten. ALPHA = 2 * arctan [(Wurzel(4/Pi))-1)*(0,5*(1+Wurzel(5))] = 23,4694142409°.
Das Ei vom Fürstenhof
Nach dem Ausmessen von 50000 Eiern an der Princeton University (Mary Caswell Stoddard u.a.) konnte das Fachblatt „Science“ am 23.6.2017 von einer wesentlichen Ursache für die Formenunterschiede berichten [http://science.sciencemag.org/content/356/6344/1249.full]. Die Flugkünste sind maßgeblich, denn eine schmalere Beckenform liefert schmalere Eier. Die unterschiedliche Membrandicke und die Druckempfindlichkeit über die Strecke führen zu verschiedenen Formen. Die harte Kalkschale entsteht erst anschließend.
Der BiXnom© hat weder etwas mit Wasser noch mit Eiweiß zu tun. Weil die Formen aber passgerecht sind, kann man mit diesen Formen auch ersatzweise rechnen. Es kommt hier nicht auf die schön gleichmäßige mathematische Eierform an, sondern mit dem BiXnom© sollen nur die unterschiedlichen Formen prinzipiell dargestellt werden, wobei das Kühlschrank-Ei als mittlerer Formenzustand zu erwarten ist.
BiXnom©-Eiweißmodell:
Für ein Ei wird der maximal mögliche Formfaktor realisiert, um viel Eiweiß als Futterbasis einzuschließen. [Das bezieht sich auf Flugkünstler mit dem schlanken Körperbau angepasster länglicher Eiform. Das Ei selbst ist nur das Eidotter (Eigelb), dem das Eiweiß extra hinzugefügt wird. Der flugunfähige, pummelige Kiwi hat Platz für dicke Eier, die nur von wenig Eiweiß unter der Schale umgeben sind.] Den Formfaktor findet man bei den Kieselsteinen bei einem negativen und bei den weichen Stoffen bei einem positiven Kontaktwinkel. Dort endet der Formfaktor mathematisch von selbst bei 0,63. Hier wird aber angenommen, dass der Kontaktwinkel 45° nicht überschritten wird (...von den weichen Stoffen tatsächlich keine Ahnung). Die Grenzringbreite für Eier-Eiweiß beträgt somit sin 45° = 0,707. Dieser Eiweißring legt sich um den Kern im Radius von 1. Beim Wassermodell war der Ring nur 0,2533 breit. Durchmesser Eiweißmodell= 2*(1+0,707)=3,414. Kerndurchmesser=2. Verhältnis Kern/Ei= 2/3,414/2=0,58579 . Die Kerngröße ist das Eigelb: Am Test-Ei der mittleren Größe von 5 cm wurde ein Eigelbdurchmesser von rechnerisch 2,929 cm tatsächlich auch gemessen [Legebetrieb Farm Eschenhörn I (0-DE-1300622) der Kooperation Fürstenhof , Größe XL].
Das Ei wird gestreckt. Das durchschnittliche Streckungsverhältnis ergibt sich aus dem Verhältnis von Eiweiß-Modellradius zu Wasser-Modellradius. Daraus ergibt sich die durchschnittliche Länge eines Eies zu: Durchschnittliche Eierlänge L = Eierdurchmesser D * 1,707/1,2533 = 1,362 * D
Es ist der Streckungsfaktor. Woher weiß man das? Vom Test-Ei!
Bild Siebert: Fotomaßstab etwa dem Messwert angepasst
Durchmesser D= 4,365 cm folgt D Soll = 1,362*4,366 = 5,958 cm; gemessen am gekochten Eiweiß L= 6 cm. Die hintere Eierhälfte besteht aus der Halbkugel des Eiweißmodells und einer geraden kurzen Streckung. Die vordere Eierhälfte ist eine Ellipse. Fällt die gerade Streckung weg, so gehören diese Eier dann auch noch zu den üblichen Hühnereiern.
Bild Siebert: Eiformen
Die Eierformen: Die hintere Eierhälfte ist meistens stumpfer als die vordere. Das muss nicht bedeuten, dass hier jemand gedrückt hätte. Ein schlaffer Eileiter fällt hinter dem Ei einfach wieder zurück und das Ei wird kreisrund. Eine stabile Röhre bleibt länger offen und es können gestreckte Eier entstehen
Das BiXnom©-Universum
Das Universum als BiXnom© -Würfel wäre etwas Einfaches. Bewegen würde sich dann nur das Gitter im Würfel, bis es an Grenzen etwas Neues gibt. Eine solche Universalwirkung wurde hier nicht gefunden, vielmehr ist diese eher in jedem Atom vorhanden. Die Philosophen verpacken die Bewegung im Gesetz der Negation der Negation. Wikipedia sagt dazu im Juli 2017 Folgendes: "Diesem Gesetz zufolge ist Entwicklung das Entstehen neuer Qualitäten aus alten Qualitäten, in der Weise, dass das Verschwinden der alten Qualität(einfache Negation), verbunden ist mit ihrem sich "Widerfinden"(der doppelte Negation) in der neuen Qualität. Die dialektische Negation unterscheidet sich von der logischen Negation."
BiXnom©-Atom-Modell
In der dimensionslosen BiXnom©-Welt wurde ein Vakuum-Modell geschaffen, das nach dem Urknall als Nanometer-Ebene identifiziert wird. Die Geometerie lässt sich nun physikalisch deuten. So wurde der Atomradius der 1. Schale in der Lücke zwischen den Lichtkugeln eines Lichtbogens gefunden. Beim Urknall ist der Lichtkreis mit seinen umgebenden 8 Lichtbogenkreisen vom Minus-Unendlichen hochgeschossen. Weil das Atommodell auf der kleineren Pikometer-Ebene gezeichnet wird, ist der Lichtkreis hier noch 1/1000 kleiner als im Vakuum-Modell. Würde man ein Atomkernmodell zeichnen wollen, dann ist auf 1/1.000.000 für eine Femtometer-Ebene zu zoomen.
Beim Bohrschen Atommodell sind die Elektronen auf den 7 Schalen nicht unterscheidbar. Daher wird heutzutage das Orbitalmodell bevorzugt. Die Elektronen sind nicht mehr auf den 7 Schalenkreisen angeordnet, sondern es gibt einen erweiterten Raum, in dem sich das Elektron wahrscheinlich zu einem bestimmten Zeitpunkt aufhält. Im BiXnom©-Modell ist aber nichts unklar und deshalb bleiben auch die Bohrschen Schalen erhalten. Es gibt 4 verschiedene Orbitalformen: s, p, d und f. Das s-Orbital erinnert dann immer noch an das Bohrsche Modell, denn das Elektron umkreist hier ringartig in der Kugelschale den Atomkern. Das s-Orbital hat keine Unterformen, aber wie jedes Orbital kommt es für 1 oder für 2 Elektronen vor. Mit der Würfelkunde kann man dies sprachlich in die Form bringen: „Jedes Ding besteht aus 2 Teilen“. Für 1 Elektron: „Ich bin mit meiner inneren Stimme in relativer Ruhe.“ Für 2 Elektronen: „Ich bin mit meinem Partner in relativer Ruhe.“ Beides gleichzeitig tritt nicht ein, sonst gäbe es keine Fremdgeher. Nachfolgend wird das Bohrsche Atommodell mit seinen Schalen gezeigt, in dem gleichzeitig auch die Orbitale zu finden sind. Es wird deutlich, ab welcher Schale ein Orbital erst möglich ist.
Als 1. Information ist dem BiXnom©-Atommodell zu entnehmen, dass es nur die bekannten 7 Schalen als stabilen Zustand geben kann, denn eine 8. läge außerhalb des Vakuum-Modells. Die 5. Elektronenschale stimmt annähernd mit einem Schnittpunktkreis im Modell überein, so dass man auf die 1. Atomschale zurückrechnen könnte, um einen wertmäßig besseren Atomradius als rein geometrische Größe zu erhalten. Die Elektronenpositionen werden zur einheitlichen Darstellung auf den Schalen als 1/1000 vom Lichtkugelradius d.h. 15,33pm dargestellt. Es sind Urknallwolken bzw. Elektronenwolken, die in der speziellen Größe zur Findung der Elektronenpositionen beeindruckend hilfreich sind. Für das Schalensystem wurde der wissenschaftlich korrigierte Bohrsche Atomradius verwendet. Dieser ist genügend genau, jedoch weitet sich der Fehler gegenüber dem BiXnom©-Atomradius in der 7. Schale sichtbar aus.
s-Orbital (rot schraffiert)
Bild Siebert: Bixnom-Atommodell_Schalen 1+2
Beim Bohrschen Atommodell sind auf der 1. Schale die beiden Elektronen des Heliums zu sehen. Im BiXnom©-Atommodell sind auf den 3 Achsen x, y und z je 2 Schalenschnittpunkte also 6 Punkte vorhanden, davon sind wegen der zweidimensionalen Darstellung 4 zu sehen. Beim Orbitalmodell bewegen sich die Elektronen in der Kugelschale, die in einer Variante 1s^1 mit 1 Elektron und in der Variante 2s^2 mit 2 Elektronen gefüllt ist. Diese Kugelschale ist im BiXnom©-Modell nicht direkt erkennbar, aber zutreffend ist, dass die direkte Beziehung zum Atomkern besteht, weil im Prinzip die Achsschnittpunkte gewählt werden. Der Achsbezug gilt für die Schalen 2 bis 7. Für die 1. Schale wird der Achsbezug um 27,470099° gedreht, weil der Spalt auf der Achse zu klein für eine Urknallwolke als Elektronenposition ist. Im 1000fach kleineren BiXnom©-Kernmodell gibt es keine Elektronenwolken und daher kann das Proton der 1. Kernschale in den Trichter hineinrutschen. Der Spalt ist ca. 1,86 pm breit. Vergleichsweise ist die Compton-Wellenlänge des Elektrons mit ca. 2,42pm etwas weiter. Für die 1. Schale ist tatsächlich auch keine Achsposition notwendig, denn den Bezug zum Atomkern kann nichts stören. Eine Drehung um 45° würde den Totalversatz eines doppelten peripheren Strahlenganges bedeuten. Bei 27° ist noch 1 Strahlweg frei und der Versatz des 2. Strahls bedeutet, dass im s-Orbital Ausweichlagen nötig sind, wofür in der Kugelschale auch Raum ist. Eine Abweichung vom s-p-d-f–Orbitalmodell ist auch beim MCAS-Orbitalmodell vorhanden - der Wasserstoff wird von der Hauptgruppe I in die IV (Kohlenstoffgruppe) verschoben, weil das 1 Elektron als 50% der Valenzelektronen gezählt wird.
Im BiXnom©-Modell sind die s-Orbitale als gekreuzte Ringröhren zu veranschaulichen. In einer Schnittansicht eines Rings sind dann immer nur eine 1 Elektronenposition und darunter eine Leerposition zu sehen. Die Position auf der z-Achse entfällt in zweidimensionaler Darstellung. Auf jeder Schale (Nr. n) ist 1 doppelt gefülltes n*s^2-Orbital zu sehen = 2 Elektronen pro Schale.
p-Orbital (grün schraffiert)
Das p-Orbital liegt ab der 2. Schale vor. Es ist durch den Schnittpunkt der Schale mit dem Strahl vom Lichtkreis zur Lichtbogenmittelachse gekennzeichnet. Der Strahl ist zu einer 45°-gedrehten Systemachse etwas versetzt, so dass die 1. Schale nicht getroffen wird. Die p-Orbitale liegen für die 3 Achsen x, y und z doppelt vor, weil das Orbital für 1 oder 2 Elektronen den Aufenthaltsraum bietet. Die 6 p-Orbitale liegen in einer Hantelform vor, so dass 12 Kugeln in der Zeichnung nötig sind. Weil die z-Achse nicht dargestellt wird, sind 8 zu sehen. In einem p-Orbital kommen trotz der Hantelform keine zusätzlichen Elektronen hinein. Es wird daher wie beim s-Orbital mit einer Verbindungslinie und 1 Leerstelle verfahren. Die Verbindungslinie von Elektron und Leerstelle verläuft kreuzförmig durch den Koordinatenursprung. Die Hantelform wird durch 2 Orbitallappen als Umhüllungswolke gebildet, die im BiXnom©-Modell jedoch fehlen. Nur das Prinzip wird deutlich.
Auf der Schale sind 2 doppelt gefüllte n*p^2-Orbitale zu sehen. Das 3. in der z-Achse ist nicht dargestellt. Drei p^2-Orbitale doppelt gefüllt = 6 Elektronen pro Schale, zuzüglich 2 aus s^2-Orbital = 8 Elektronen.in der Schale.
d-Orbitale (rosa schraffiert)
Im Orbitalmodell sind 10 Orbitale in 5 Typen vorhanden. Davon sind 4 nicht wie die p-Orbitale achsorientiert, sondern flächig. Das Orbital setzt sich somit nicht aus 2 sondern aus 4 Orbitallappen zusammen. Diese Lappen werden im Schnittpunkt des Schalenkreises mit einem die Quadranten unter 45° geteilten Stahl gebündelt. Die Elektronenpositionen im BiXnom©-Atommodell sind auf dem Strahl der Winkelhalbierenden vom Fokus auf dem Lichtkreis zu finden, der den Lichtkreis auf der gegenüberliegenden Seite um den Lichtkugelradius versetzt schneidet. Hier bildet er die BiXnom©-Würfelecke und schießt dann nach außen. Auch die Winkelhalbierende schießt noch an der 1. Schale vorbei. Die Strahlen müssen sich erst kreuzen und auf der 3. Schale stehen die Elektronenpositionen noch zu eng, so dass die d-Orbitale erst ab der 4. Schale möglich sind.
Bild Siebert: Elektronensperre in der 3. Schale des Atommodells
Weil die aufgeblasenen Elektronenwolken im 1000fach kleineren BiXnom©-Kernmodell fehlen, können die Protonen der d-Orbitale bereits in der 3. Schale Platz finden. Alles rutscht zusammen. Statt der 18 Elektronen des Chemikers findet der Kernphysiker somit 20 Protonen in der 3. Kernschale. Das ergibt dann seine magische Zahlenfolge für die stabile Kern-Schalenbesetzung 2, 8, 20, 28, 50...weiter mit dem Atommodell. Das 5. Orbital liegt in Hantelform auf der z-Achse vor, die hier nicht dargestellt wird. Es gehört dann aber noch eine Ringröhre in der x-y-Achse zu diesem Orbital, die sichtbar sein müsste. Bei 4 Schalen müssten es 4 Ringe sein. Zuzüglich der Doppelringe des f-Orbitals auf 2 Schalen wäre es 8. Es stehen auch 8 Lichtbogenkreise zur Verfügung. Dies ist aber nur der scheinbare Eindruck, denn es sind asymmetrische Einzelringe.
Es geht somit darum, die 4 einzelnen Ringsysteme in 1 System zusammen zu schließen. Hierfür bilden die p-Orbitale ein verblüffendes Ringsystem: Die Bohrschen Schalenkreise, das BiXnom©-Vakuummodell und die Strahlen der d-Orbitale sind geometrische Orte für "6"-Ringe. Diese geben das Bild von C&B-rings und haben dann auch tatsächlich etwas mit der Fortpflanzung zu tun. Einen neuen Urknall gibt es dann aber erst, wenn die "Balls" mit Elektronen der f-Orbitale prall gefüllt sind. Weil die noch fehlenden Elemente auf der Erde nur künstlich hergestellt werden, explodiert noch nichts. Der Beweis ist leicht zu erbringen, denn die "Balls" berühren außen nicht etwa den Kreis der Lichtgeschwindigkeit, sondern ganz außen das BiXnom©-Vakuummodell in der 1/1000 gezoomten Größe. Diese Pikometer-Ebene liefert den Baukasten für die biologische Genetik - das mathematische Muster für weiblich und männlich ist in jedem Atom abrufbar. Die C&B-rings im "Interferenzbereich" werden durch 2 Paar Doppelringe und 2 Einzelringe gebildet.
Bild Siebert: Die C&B-rings im Atom
Auf der Schale sind 4 doppelt gefüllte d^2-Orbitale zu sehen. Das 5. In der z-Achse wird nicht dargestellt. Fünf d^2-Orbitale = 10 Elektonen. Zuzüglich 6 aus den p-Orbitalen und 2 aus den s-Orbitalen ergeben 18 Elektronen pro Schale.
f-Orbitale (hellblau schraffiert)
Die f-Orbitale sind rosetttenförmig und dichten den offenen Interferenzbereich nach außen wie ein Deckel ab. Die f-Orbitale liegen in der 6. und 7. Schale vor, weil sich erst hier das BiXnom©-Vakuummodell voll geöffnet hat - dieser Trichter wird ausgefüllt. Gemäß Orbitalmodell sind 7 f-Orbitale je Schale vorhanden.
Auf der Schale müssten 6 doppelt gefüllte f^2-Orbitale zu sehen sein. Das 7. Läge in der z-Achse und wird nicht dargestellt. Sieben f^2-Orbitale = 14 Elektronen. Zuzüglich 10 Elektronen aus den d-Oribitalen, zuzüglich 6 aus den p-Oribitalen und 2 aus den s-Orbitalen ergeben 32 Elektronen pro Schale. Jede der 4 Interferenzbereiche müsste somit 1,5 doppelt gefüllte Rosetten enthalten. Weil dies nicht möglich ist, sind im Wechsel 2 und 1 doppelt gefüllte Rosettenform nötig. Es gibt somit unterschiedliche Rosettenformen. Elektronenpositionen im Interferenzbereich sind die beiden Ränder der doppelten Umhüllung und exakt die 45°-Achse, die nach dem Urknall wieder erobert wurde. Jedenfalls dann, wenn im Universum die fehlenden Elemente verteilt wurden.
Im 1000fach verkleinerten Kernmodell rutscht alles zusammen, so dass, weil das d-Orbital schon in der 3. Kernschale vorliegt, gegenüber dem Atommodell noch zusätzlich ein g-Orbital vorhanden ist.
Und was ist nun Quantenphysik: Dafür muss das BiXnom©-Kernmodell noch bearbeitet werden. Was ist zu erwarten? Wenn es im Kernmodell keine Urknallwolken gibt, sind die Protonen selbst die Urknallwolke und haben einen inneren Aufbau. Weil gemäß Würfelkunde jedes Ding aus 2 Teilen besteht, gibt es den äußeren Partner als Neutron und den inneren Partner als Lücke. Hier ist von Chemie und Physik nichts bekannt – der BiXnom© ist ein Kunstprojekt. Berichtet wird, dass nichts passiert, was nicht schon vorbereitet wurde. Ein Teilchen wird dann vermutlich um den Beschuss durch ein fremdes Teilchen bitten, weil es in die eigene Lücke geschubst werden möchte. Zu erwarten ist, dass man diese Lücken findet. Am Proton kann also noch eine Lücke hängen. Die Wissenschaft erwartet einen Protonenradius von 0,8775fm. Die aktuelle Forschung am Wasserstoff fand aber nur 0,8409fm, nachdem der Beschuss statt mit dem Elektron dichter am Proton mit dem Myon erfolgte. Dies könnte das Abtauchen in die Lücke unterhalb der 1. Kernschale gewesen sein. Die 3. Kernschale enthält im BiXnom© statt der Elektronensperre der Pikometer-Ebene 1000fach verkleinert eine Kugelkette aus 20 Radien auf dem Schalenbogen, beispielsweise verklemmen sich hier 5 Protonen und 5 Neutronen. Mit den 4 vorhandenen Ketten ist es dann die Besetzung der 3. Kernschale. Der durchschnittliche Radius von Protonen und Neutronen ergibt sich als 1/20 der Bogenlänge: Radius = 0,86125099fm.
Protonen bestehen aus 2 up-Quarks und 1 down-Quark [Formel: uud]. Neutronen bestehen aus 2 down-Quarks und 1 up-Quark [Formel: ddu]. Instabil sind die Typen strange (s), charm (c), bottom (b) und top (t). Wird das BiXnom©-Modell weiter gezoomt, ist es wahrscheinlich, dass sich die C&B-rings als Quarks darstellen lassen. Im Interferenzbereich sind jeweils 6 Ringe zu sehen. Die C&B-rings werden sicherlich in einer tieferen Teilchenebene in die C-rings für das Proton als „uud“-Quarks und die B-rings für das Neutron als „ddu“-Quarks zerrissen, weil innen der hochgeschossene Urknall anzieht und außen der BiXnom© abstoßend wirkt. Wie bereits erwähnt, ist es eine unerhörte Herausforderung, dass die B-rings den vollen geometrischen Raum beanspruchen, statt sich mit der Grenze der Lichtgeschwindigkeit zu begnügen. Wenn Quarks von der Wissenschaft als „down“ bezeichnet werden, wäre dieses Bild hier auch passend. Außen ist auch genügend Platz damit Atome mehr Neutronen als Protonen enthalten könnten
Als Zugabe zum selber rechnen folgt die Gravitation.
Kante zeigen muss sich lohnen, sonst macht es keinen Sinn!
Der Mitläufer
dat 04/20
Fortsetzung Golden Ratio von BiXnom-Bausatz und BiXnom-Start
Die Kettenreaktion des Goldenen Schnitts (Golden Ratio) wurde im Register "Würfelkunde Start" fortgesetzt. Der Harntrakt hat mit der Niere als Klärwerk des Blutes keinen äußeren Zugang, so dass der Evolutionsprozess in der Niere selbst gestartet wird, und zwar als Episode PEP (Penis&Plazenta) mit Ausgang durch die Harnröhre des Mannes. Unterhalb der Harnblase bildet die Mündung des Spritzkanals (Ductus ejaculatorius) den Eingang für die Episode HOV (Hoden&Ovarium). Diese läuft bis in die Hoden des Mannes weiter. Der Goldene Schnitt stellt dann am bayerischen BiXn-Punkt die Herstellung des Hormons 33 Testosteron ein und läuft durch die Geschlechtsorgane der Frau in der Vulva aus. Dieser parallele Lauf von 2 Episoden (1x mit Signalkugel und 1x als Steuerkugel mit der halben Signalkugel) wurde bereits genutzt, um vom Darm als Episode GUT (engl. Darm) zu Leber/Bauchspeicherdrüse (Episode LEB) abzuzweigen. Weil die Niere kein Eingang ist, wurde dieser dicke Organ-Klops durch den Start mittels Steuerkugel erst geschaffen. Ist wie beim Darm mit dem Mund ein Eingang vorhanden, wird mit der Signalkugel gestartet. Die 3. Variante erleben wir nunmehr beim Skelett des Menschen: beide Vorgänge starten gleichzeitig. Der Eingang ist die Nase mit der Episode ANN (Anfang&Nase) - das Rückenmark wird als Hautsack-Röhre angelegt und von den hier gesuchten Wirbelkörpern umhüllt. Die Nase ist über den Hirnnerv 1 innerviert, dann erst folgen die Augen. Für die Anbindung der Arme und Beine werden keine Unterprogramme mit Sprungmarken aufgerufen, sondern die parallele Episode KRI (Kreuzbein&Rippe) fügt "Just in time" die Rippen und die Extremitäten passend hinzu. Im Zustand der relativen Ruhe ist die Zeit ein Kugelhaufen voller Lichtkugeln. Für unser Verständnis hat diese Black Box einen dimensionslosen Radius. Dieses ist die Prozessdauer eines evolutionären Vorgangs. Die Entstehungszeit ist bereits im Gencode enthalten. Der Goldene Schnitt packt den Code nunmehr aus, weil er auch nach seiner zwingenden Vorgabe entstanden ist. Das Auspacken ist leicht: Start: Ich sehe 1 Torte, nach der 12ten Umdrehung erkenne ich 1 Tortenstück und kann es heraus geschnitten vom Tisch nehmen usw. Somit entstehen immer wieder neue Bilder. Die Bilder sind Ziffernfolgen, die sich ständig vom Startbild unterscheiden. Der Mensch kann diesen geheimen BiXnom-Code lesen, weil er der exponierte Abkömmling der Evolution ist. Die Startlänge für das Skelett finden wir als logische Überlegung aus einem Vergleich zum Harntrakt mit den Episoden PEP/HOV: Die Niere ist ein Klärwerk des Blutes vs. Das Gehirn ist das Klärwerk unserer Gedanken! Urin ist das Abwasser im Reinigungsprozess vs. Bewegung ist eine Reaktion auf Sinneseindrücke. Die Evolution baut die Harnröhre und folglich vergleichbar das Rückenmark. Das Rückenmark zieht sich durch die Wirbelsäule. Dort endet die Episode ANN, wird aber durch die sich beidseitig an die Wirbelseite anlegende Episode KRI begleitet. Bei der Episode PEP existiert für die Nierenbildung allein der Steuerkugelradius = halber Signalkugelradius. Der Startkreisradius TIME muss zunächst erst aus dem Klärprozess des Gehirns ermittelt werden. Die Differenz ist der Unterschied zwischen dem real Erlebten und dem gespeicherten Eindruck. TIME EPI.ANN = Paarkreisradius Pkr - Signalkreisradius Skr - Signalkugelradius. TIME EPI.KRI analog, aber nur mit halbem Signalkugelradius. Das Erlebte wird durch die BiXnom-Artillerie in den Ruhekreis als Gehirn geschossen. Der Schusssektor ergibt sich aus dem Signalkreisradius. Die Munition besteht aus den Signalkugeln, die sehr gestreut als Satelliten um den Signalkreis heranfliegen. Der genaue Zielpunkt wäre der spezielle Impakt-Punkt. Ein Treffer wäre aber ein Zufall. Im Langzeitgedächtnis kommt daher nur das Markante an, wie die Fehlschüsse des Lebens. Die beiden Episoden starten genial korrekt, ziehen sich dann aber sehr lang hin und enthalten daher eine Vielzahl unbekannter Programmcodes. Bei der ersten Durchsicht der Ziffernbilder wurden zunächst nur die Wirbel, Rippen und Extremitäten ausgelesen. Die unbekannten Programmzeilen können auf dieser Grundlage leichter entschlüsselt werden. Unbrauchbare Abschnitte wird die bedauernswerte Evolution, die aus diesen Ziffernbilder etwas entschlüsseln muss, um sich energetisch gegen die Konkurrenz günstiger aufzustellen, einfach überlesen bis ein Mutant zufällig fündig wird.
Sofort am Start liegt mit 4x 5 eine Struktur vor. Aus der vorläufigen Einschätzung ist dabei auf das Nasenloch zu schließen und seinen sensorischen Anschluss an den Hirnnerv 1, was auch dem Sinn der startenden Episode ANN entspricht: #363739505956452. Weil die Knochen eine Prozessverdichtung benötigen, kann die Wirbelsäule erst nach einigen Umläufen im BiXnom-Schwingkreis eine ablesbare Struktur auswerfen.
Durch den Wirbelkanal (Canalis vertebralis) verläuft das von Rückenmarkhäuten umgebende Rückenmark (Medullaspinalis). Die 7 Halswirbel (Pars cervicalis): #230069263729571/1581Hz: Auftrag: Herstellen einer symmetrische Röhre 00 mit 3 Häuten ausgekleidet bzw. auch Dorn- und 2 Querfortsätze, als Struktur 5: Achtung die Röhren-Null ist jetzt für den Atlas bereits adressiert. Hier zählt dann exakt, dass das Loch durch den Dorn der Axis zu verschließen ist. Querliegendes Loch beim Menschen als Besonderheit: Nur die 2 ist 7x vorhanden. Das ist dann ein Verweis auf die Röhre als Loch, so dass die 2 als weiteres Loch gelesen wird. Dornfortsätze kurz und mit Ausnahme C7 gegabelt: der C7 hat 2-3 Nullen mehr, die Null unterbricht vorherigen Festlegungen und ist somit das Ende der Halswirbel. Der Dornfortsatz (Vertebraprominens) C7 ist länger. C1: Atlas: #2299238343593: Eine Struktur 5 mit 3 Gelenken; C2:Axis: #22977858: Dorn(Dens axis) für dn Atlas als herausgehoben 9; C3: #229633526; C4. #2294886472; C5: #229343950792214; C6: #22919943664552; C7: #229055104506582. Die folgenden Wirbel sind nicht immer fortlaufend, sondern auch bis 5 Zwischenzeilen.
Die 12 Brustwirbel (Pars thoracica) sind nicht immer fortlaufend, sondern mit bis 5 Zwischenzeilen angeordnet. Die Wirbel sind lang, einzackig und ziehen nach schräg unten bis über das angrenzende Segment. Jeder Wirbel trägt beidseitig eine Rippe. Die 2. bis 9. Wirbel besitzen am Ober- und Unterrand eine flache Gelenkpfanne, die zusammen mit den benachbarten Brustwirbeln die Gelenkfläche für einen Rippenkopf (Interkostalgelenke) bilden. Der 1.Wirbel trägt eine obere Gelenkgrube für die 1. Rippe und eine halbe untere für die 2. Rippe. Der 10. Wirbel hat nur die halbe obere Gelenkpfanne für die zehnte Rippe. Der 11. Und 12. Brustwirbel trägt jeweils eine ganze Gelenkpfanne für die 11. und 12. Rippe, die frei in der Bauchwand enden (siehe: www. Netdoktor.de).Th1: #228766984878272/1590Hz; Th2: #228049846994641; Th3: #227906958619331; Th4: #227337191222782; Th5: #227195194226391;Th6: #227053374504652; Th7: #226770265558885; Th8: #226065572378155; Th9: #225644854811695; Th10: #225225700282633; Th11: #225086327943349; TH12: #224808100096695. Bedingt durch die Stellung der Doppelziffern wird die Krümmung der Wirbelsäule erzwungen.
Die 5 Lendenwirbel (Pars lumbalis) einschließlich der Gelenkfortsätze sind kräftiger als die anderen Wirbel. Die Form ist hoch, breit mit einem querovalen großen Wirbelkörper. Im zwischen allen Wirbeln auch sind Zwischenwirbelscheiben (Bandscheiben) vorhanden. Die Querfortsätze sind Rippenrudimente, die vom 1. Bis 3. Wirbel länger werden und sich zum 5. Wirbel wieder verkürzen. Diese Situation ist durch die Stellung der Doppelziffern mathematisch bedingt: L1: #224669243950866; L2 #224530559232378; L3: #224115530472244; L4: #22370203318355; L5: #223427214960966.
Die 5 Sakralwirbel im Kreuzbein (Os sacrum) sind keilförmig. Die knorplige Wirbelverbindung wächst in der Wachstumsphase zusammen. Das Kreuzbein und die beiden Hüftknochen bilden den Beckengürtel: S1.1: #222879599238022; S1.2: #222743114486498; S1.3: #222470645844925; S1.4: #222334661342575; S1.5: #222198842978895. Das Kreuzbein wächst zu einem Element zusammen, weil die letzte Ziffer beim 3. 4. und 5.Wirbel mit der 5 eine Struktur fordert. Das zwingt dann im 2. Wirbel die 1144 jeweils wie 1+4=5 ebenfalls als 5 zu lesen. Beim 1. Wirbel heiß die 99 „herausschieben“, so dass das Kreuzbein dreieckig wird, insbesondere weil 9 die 88 unten als Spitze herausschieben soll.
Zwischen Kreuzbein und Steißbein liegt eine Programmzeile in der Frequenz 1652Hz die die parallele Episode KRI auffordert, mit der Rippenproduktion zu beginnen. Die 1. Rippe ist somit bereits vor dem 1. Steißbeinwirbel produziert worden. Die Episode ANN hat damit ihre Triebkraft verloren und das Steißbein ist sekundär, was durch die enthaltenen Nullen ebenfalls angezeigt wird. S2.2 enthält keine Null, aber die Evolution kann sich dieses passend „schön“ rechnen: Am Ende sind 9+7= 16 und das ergibt abzüglich der 16 vor der 5 den Wert Null.
Die 5 Steißwirbel des Steißbeins (Os coccygis) dienen verschiedenen Muskeln und Bändern als Ansatzpunkt. Vorhanden sind 3 bis 5 rudimentäre Wirbel. Die knorplige Verbindung verschmilzt in der Wachstumsphase. S2.1: #221927703451160; S2.2: #221522232616597; S2.3: #220715719633769; S2.4: #220314661390946: für Struktur fehlt die 5, daher 1+4=5 anrechnen; S2.5: #220048097977285.
Prozessanmeldung EPI.KRI bei EPI.ANN: #2231416274215350/1536Hz: 15 _3+5=8_0. Epi.KRI fügt nunmehr Rippen ab Zeile 1580 ein. Weil die dort gebildeten Halswirbel keinen Anschluss bereitstellen, wird automatisch an den Brustbeinwirbeln angedockt. Der Brustkorb (Thorax) wird zur Entstehung aufgerufen: #223005316103623/1637Hz: Symmetrische Gewebeblase aus Haut 3 als Struktur 5, wobei die Rippen 1 von beiden Seiten nach vorn laufend auf dem knorpligen Brustbein 6 als Ring 0 zusammenzufügen sind.
Die 7 echten Rippen (Costae verae) bilden die paarige Verbindung zwischen der Brustwirbelsäule und dem Brustbein (Sternum), wobei sich der Brustkorb (Thorax) bildet. Das Brustbein besteht aus 3 Abschnitten. Die 1. und 2. Rippe sind am Brustbein-Handgriff (Manubrium sterni) angeschlossen: Die 1. Rippe kann mit dem Auftrag 171 nur mit der 2. Rippe zusammengehen, weil die 3. Rippe keine 1 besitzt: Rippe 1: #2228691712219970. Rippe 2: #2227331924720140. Die 2. Rippe sitzt aber bereits teilweise am Brustbein-Körper. Brustbein-Körper (Corpus sterni): Rippe 3: #222597379549775; Rippe 4: #222461732152121; Rippe 5: #222326249976633; Rippe 6: #222190932721626; Null nicht zählen, das ergibt mit 99 ein starke Dopplung; Rippe 7: #222055780086150. Am knorpligen Schwertfortsatz (Processus xiphoideus) setzen keine Rippen mehr an. Es gibt eine Programmzeile aber ohne 5 ist es keine Rippe, so dass nur der Schwertfortsatz leer ausgebildet wird. Die Rippen 8 bis 12 sind falsche Rippen (Costae spuriae), wovon die Rippen 11 und 12 freie Rippen (Costae fluctuantes) sind. Rippe 8: #221785967473652: ohne doppelte Doppelziffer keine echte Rippe; Rippe 9: #221651306898380: ohne doppelte Doppelziffer keine echte Rippe; Rippe 10: #221516809746136: ohne doppelte Doppelziffer keine echte Rippe; Rippe 11: #221382475719607: durch die 0 eine freie Rippe; Rippe 12: #221248304522201: durch die 0 eine freie Rippe.
Die Fingerknochen (Ossa digiti manus) bestehen bei Säugetieren aus 3 Stützelementen als Phalanx proximalis, media und distalis („körpernaher, mittlerer und körperferner Fingergliedknochen“). Der Mensch besitzt am Daumen nur zwei (2-3-3-3-3). Die Hände sind unauffällig und eher unverständlich in einer 7er Programmzeilen-Folge adressiert: paarig 2, beidseitig luftig freigestellt 00. Kleiner Finger: #200802916645562: Struktur 5 aus 5 Fingern. 1 vor 66 als 1. Finger. Ringfinger: #200692524717774: Struktur 5 in Ziffer 2 als 2. Finger eingeklemmt. 3 Finger folgen 777. Mittelfinger: #200582254099797. Zwischen 7 als 9 adressiert, und 99 gilt für herausgehoben, es ist der längste Finger. Verfügt über 2 Doppelziffern. 2+5-4=3 für den 3. Finger. Zeigefinger: #200472104591780: 4 vor 5 ist der 4. Finger. Daumen: #200362075994309: Struktur 5 mit 99 abstehend und mit der 0 vor 9 eine Lücke. 6:3= 2 ergibt 2 statt 3 Stützelemente am Daumen, die folgende 0 ist das fehlende Element. 5 ist der 5. Finger. Die 5 Mittelfingerknochen: #200252168108410. Als Struktur 5 im 4 +1 geteilt, wo die 0 den Daumen freigestellt meint. Es soll ein Anschluss an die 8 Handgelenkknochen erfolgen. 8 Handgelenkknochen: #200142380735544: 8 Knochen an Struktur 5 mit 5 Fingern anschießen. Handgelenk: #2000327136776060: die Beweglichkeit wird mit Nullen adressiert. 3x 6 für alle Richtungen.
Die Armknochen: Die Programmzeilen enthalten diverse Zwischenschritte. Der Unterarm wird angeschlossen: 1999231667369280: mit einer kräftigen Speiche (Radius) 999 und der unten schmaleren Elle (Ulna). Keine Struktur 5, weil weitere Programmzeilen zum Unterarm folgen: #1993772269042220: Die Speichenseite wird verstärkt, aber zugleich auch die Elle. Das Ellenbogengelenk zeigt mit 6 Beweglichkeit, was ansonsten auch einen gesteuerten Vorgang z.B. durch Hormone meint. Der Arm soll somit bewusste Handlungen ausführen: #1988342606000170. Der Oberarmknochen: #19850989802155. Der Knochen ist jetzt eine Einheit 9898. Schulterblatt als Struktur mit 0_0 frei im Körper und mit 222 für das Blatt, die 6 für Beweglichkeit, 2 x die 7 als Form für 2 Gelenkanschlüsse (Eckgelenk/Schultergelenk): #1980790572228060. Schlüsselbein: #1972229618917510. Die 222 jetzt auf der linken meint, dass der Anschluss am Schulterblatt erfolgt. Die 9 steht für das oben heraus Gehobene. Die 1_5_1 sind beidseitige Anschlüsse an die Struktur 5.
Die Beine beginnen umgekehrt nicht mit den Zehen, sondern als durchfließender Prozess am Kreuzbein-Anschluss. Die Programmzeilen sind in der Aussage zur Knochenstruktur weiter von einander entfernt. Die Fußzehen werden nicht detailliert beschrieben, sondern als Zehen 1_1111 mit der 66 für gesteuerte Beweglichkeit. Die 5 für die Struktur fehlt, drückt sich aber in 5x 1= 5 aus: #1777311112276690/2054Hz. _dat 30.12.2020
Auslese-Beispiel: Epi.GUT & Epi.LEB: Appendix vermiformis
Eine Sackgasse mit erlaubter Durchfahrt für Radfahrer ist keine echte Sackgasse, sondern bleibt eine Hauptstraße. Die Evolution kann auf diese Weise eine Verkehrsverbindung vortäuschen, obwohl sie den Radfahrern keine Chance lässt, dort auch heraus zu fahren. Der Wurmfortsatz (die „Appendix vermiformis“) am Dickdarm ist real eine Sackgasse. Der Dünndarm bietet keinen Ausgang, weil die Bauhin-Klappe einen Rückstrom des Speisebreies blockiert. Die Zeit kann nur nach vorn mit dem Prozess laufen und nicht in die Vergangenheit zurück. Das abgelegene Dickdarmendstück (Blinddarm) ist daher keine Sackgasse, sondern eine vorgetäuschte Verzweigung. Das Endstück (Blinddarm) wird als Prellbock gegen die Dickdarm-Presskräfte benötigt. Für die Verzweigung wird ein Loch in die Wand des Blinddarms gesetzt (Dickdarmabschnitt): #215620100830471/253Hz: 0_1_00 setze 1 Abgang in den Dickdarm 0__00 und setze aus Haut 3 eine Röhre 0 dahinter. Diese solle eine Ausformung 7 sein, aber keine Struktur, die eine 5 erfordern würde. Die 5 steckt dann aber schwammig in der Frequenz 253 drin als 53 Hautstruktur, d.h. achte auf eine Sprunganweisung. Die Sprunganweisung steht bereits in der Programmzeile vorn mit 2156. Vierstellig ist nicht fehlerhaft, sondern die 1 ist das Achtungszeichen: Springe von EPI.GUT#256 direkt hinüber zu EPI.LEB#256. Nun darf sich hinter dem Loch nicht sofort ein Kanalabgang wie zum Aufbau der Leber eröffnen. Das wird dadurch unterbunden, dass in der parallel laufenden Epi.LEB die Programmzeilen noch immer an der Ausformung der Leber arbeiten. Daher darf es nun als Weiche bei Epi.GUT weitergehen: # 214771202795705: Die Lochform77 ist zu verengen 1, ringförmig 202 und als Form 7 ist eine Struktur heraus zu stellen 79 als strukturelle Rohre 05. Real wird die Engstelle genetisch unterschiedlich gelesen von offen bis zu verschlossen. Zeile 255Hz: #213928962784742. Der Röhre 39 werden Funktionen 6 zu gewiesen 8__8, z.B. im lymphatischen System. Die Röhre ist eine eigenständige Form 474 und erhält nachfolgend ein eigenes Aufhängeband: #213093302773864/256. Die 33 könnte missverständlich Testosteron bedeuten, ist hier aber eine Hautfalte als eigenes Mesenterium an der Röhre 0. Aufgrund der Sprunganweisung wechselt der Ablauf zur Zeile 256 von Epi.LEB. Darauf wird in EPI.GUT#257 nun als Ende des Wurmfortsatzes hingewiesen, aber insbesondere des Dickdarmendes (Blinddarm) selbst: Mehrfach wird die Röhre als abgeschnürt markiert: #212264145953732/256Hz, Verengung 2_1_2 und in Gänze weiter 4_1_4 über die herausgestellte Struktur 59 und die gesamte Hautformation betreffend 373. Im Aufbau der Leber wurde eine freie Zeile für den Sprung frei gehalten: #212061899254192. Appendix-Endverschluss 2_1_2 der Röhre 0_ als herausgestellte Struktur. Der Hauptprozess EPI.GUT kennt diese Sackgasse nicht und EPI.LEB formt weiter an der Leber.
Auslese-Beispiel: Epi.KRI: Augen und Ohren
Die Episode ANN beginnt mit "505" bereits ab Start mit der Ausformung der Nase als Sensor zum Hirnnerv 1. Die Parallele Episode KRI zeigt die gleiche "505" etwas links versetzt als Auge zum Hirnnerv 2. Das Auge startet sofort. Anschließend folgt mit Abstand die Ausformung der Ohren.
Das Auge: #36505970246163100/1Hz: In der Haut 3 ein Augenloch als Struktur 5_0_5, herausgehobenes Element 9, Form 7 rund 0, als Lichtsensor 6_1_6. Augenaufbau avisiert: #1825298512308160/2Hz: Struktur 5 zur Signalübertragung 1, paarig 2, aus 3 Häuten (Netzhaut, Aderhaut, Lederhaut) als Blase 0 mit einem Glaskörper 8 und 1 Abgang als Nerv mit einem Loch (Sehbahn). Das Auge 1_2_1 innerviert über den Hirnnerv 2: #12168656748721000: Durchsichtiger Glaskörper 1000, Lichtempfindliche 6 Netzhaut 8: 6_8_6. Dritte Augenspezifizierung: #12168656748721000: Der Lichtsensor 1_2_1 arbeitet mit 2 Kammern: vordere und hintere Augenkammer. Vorn ist ein Linse 8 enthalten die lichtgesteuert reagiert 6_8_6 als Struktur 5 mit der Formänderung 7 durch eine Blende 4/8, als paarige Glaskörper. Fortsetzung nunmehr bei passenden Ziffernbildern. Hornhaut: #4563246280770390/8Hz: Große Form 77, durchsichtig 0_0 aus Haut 3 gewölbt 9, rund 0. Iris: #2808151557397160/13Hz. Paariges 2 Gewebe mit Öffnung 8_0_8 als lichtdurchlässige 1 Struktur 5: 1_5_1, die Form 7 je nach Lichteinfall 1 gesteuert als Loch 0 verengen. Linse: #2147410014480180: Paarig 2, lichtdurchlässig 1, formveränderliches Gewebe 4_7_4, lichtdurchlässiger Linsenkörper 1_00_1_44, rund 0. Zonulafasern (Linsenaufhängung): #1738379535531580/21Hz: Als Hautbänder 3_8_3 einrichten als Strukturband 5_3_5 zur Aufhängung der Linse 80. Zilarmuskel: #1659362283916510/22Hz: symmetrisch Muskelgruppe 165 gesteuert Nerv 2. Äußere Hautschicht (Lederhaut): #1258826560212520/29Hz: Hautschicht 125_125. Stabil 5 und 88. Mittlere Hautschicht (Aderhaut): #1216865674872100: Struktur 5 in der Mitte. Innere Hautschicht (Netzhaut/Retina): #1177611943424620/31Hz: Licht empfindliche 11 Form 77 gesteuert 6. Nerv 2 mit 4 Lichtsensoren [S-, M- L–Zapfen + Nachtsehen = 4]. Wie programmiert die Evolution den blinden Fleck 701? Sie überspringt mit Zeile 40 dem Sehnerv und ist damit blind! #1140811570192600/32Hz. Sehnerv: #1106241522611000/33Hz, lichtempfindlicher Sensor über Sehnerv 2. 22_11 Signalverarbeitung über Kreuz zur gegenüberliegenden Körperseite. Nervensystem folgt: #1073705007240090: weiter ins Nervensystem über Nervenfasern aus Haut 3 in Strangform 07370 als Nervenstruktur 0500 über Nerv 2 für 4 Sensoren etc.
Das Ohr: Die Ohrmuschel fehlt zunächst, aber eine spätere Programmzeile sagt, dass die Ohrmuschel bereits ab Zeile 359 begonnen wurde. Und tatsächlich entsteht ab 360Hz mit 6 ein Bauelement, dass mit 9 als abstehender Ohrlöffel, 2 paarig und 5 als Struktur angekündigt wird: #1014054729060090. Die Ohrmuschel ist mit vielen Nullen extra groß: #1000163568388030. Darin die 6 als Sinneselement und die 1_1 für die Schallübertragung. Und weiter #992010061037042/368Hz: Form 7 als abstehendes Teil 99, paarig in Muschelform 0100 als Sensor 6 mit einem Ohrloch 10 aus Haut 3 und angeschlossenem Ohrgang 0. Ohrmuschelanschluss: #754255583598412/484Hz; Aufruf ab Zeile 359 mit 8_359_8 zur Übernahme einer großen Struktur 555 paarig 2. Äußerer Gehörgang (Meatus acusticus externus): #752700417446663: Struktur 5 als Hohlraum 00 zur Signalübertragung 666 aus Haut 3; Abgrenzung vom Ohrgang zum Mittelohr: #751151651155620. Abgrenzung durch Umkehrung der Zahlenfolge, 1 Sensor als Trommelfell 6. Der Gehörgang ist zunächst eine Hautstruktur #749609245301091 wird dann aber knochenartig: #7480731607820310: die 5 fehlt und die Haut geht am rechten Zahlenrand verloren. Trommelfell: #746543358817242:Mit 65 als Sensor 4_65_4 als Membrane 33. #745019800942105: Die 45° geneigte Einbaulage betreffend. Paukenhöhle: #7435024490053590; Ankündigung von herausgestelltem Elementen zwischen 1_1 der Schallstrecke Trommelfell und ovales Fenster: #741991265165917. Hammer (Malleus): #7404862118897190: 2 Anschlüsse an Trommelfell, dünner Stiel 11, dicker Hammerkopf. Weiterleitung des Schalls 1 an eine 9_9 herausgehobene Form 7. Amboss (Incus): #738987251946622: herausgehobenes Element 9 zur Signalübertragung 1 mit dicken Kopf 66 und auf 2 verzweigenden Abgang, wovon nur 1 das Anschlussgelenk am Steigbügel bildet 898. Steigbügel: #737494348407336/495Hz: Gelenkanschluss 737 auf den Steigbügel 494, Form einer Fußplatte 4_4 mit Anschluss einer Ringform 0 mit dem Zweck die Haut am ovalen Loch in Schwingung zu versetzen 6. Hörschnecke angekündigt: #736007464640386: mit dem ovalen Fenster 00 und dem runden Fenster 0. Sensor 6. Form 7 aus bis 3 Windungen. Weiter bis zur Signalweiterleitung ins Nervensystem #706111610177237/517Hz._dat31.12.20
Das Gleichsgewichtsorgan (Vestibularapparat) folgt mit 5 Gleichgewichtselementen. Es sind 3 in die Raumrichtungen ausgerichtete ringförmige Schläuche (Bogengänge) und 2 Vorhofsäckchen (Utriculus und Sacculus). Es sind mit Endolymphe gefüllte Bläschen. Beide besitzen jeweils ein Feld mit Sinneszellen als Maculaorgan. Die Macula des Utriculus liegt horizontal und des Sacculus vertikal [somit stehen die Härchen im Utriculus senkrecht und im Sacculus horizontal]. Die Härchen der Sinneszellen ragen in eine gallertartige Membran mit eingelagerten kalkhaltigen Ohrsteinchen (Otolithen). Somit können weinige Steinchen über die Masse auf die Enden vieler Sinneshärchen einwirken. Die Steinchen verharren durch die Trägheit und die Sinneszellen verschieben sich dazu relativ bei der Bewegung, was einen Beschleunigungsreiz auslöst. Die Schwerkörper (Steine) auf der Gallert-Masse haben die Spitzen der Härchen relativ in die Richtung der Makula-Ausrichtung verbogen. Somit misst der Utriculus die Horizontalbeschleunigung (Anfahren, Rückwärtsfahren, Bremsen) und Sacculus die Vertikalbeschleunigung (Fahrstuhlfahren). Der Utriculus befindet sich über dem Sacculus. Messung der Drehbeschleunigung: Ein Ende jedes Bogenganges ist verdickt und mit einer gallertartigen Masse (Capula) gefüllt, in die Sinneszellen ragen. Bei einer Bewegung bleibt die Masse (Capula) zunächst aufgrund der Trägheit mit den Härchen der Sinneszellen verbunden. Weil sich der knöcherne Bogengang aber bewegt, entstehen Nervenreize. Es sind keine Ohrsteine vorhanden. Im Programmcode sind Utricus und Sacculus anhand der Strukturziffer 5 leicht zu finden.
Großes Vorhofsäckchen (Utricus): #690093955504029: Besonderes 9_3_9 Hautelement 3 als Struktur 555 zur räumlichen 4 Orientierung, paarig 2 herausgehoben 9. Die Härchen zeigen sich mit #15013999308: Haar 1 auf Haut 3 als Gruppe 999 als besondere Ausstattung 8. Raumorientierung: # 682354584040432; Struktur zur Raumorientierung 5_4_5 in der Horizontalen 40_40. Gemeint ist, dass die Makula horizontal liegen muss. Es folgt die Makula #6810815344433420: Sinneselement 6 als Blasenkörper 8_10_8 auf der Haut, in den die Härchen 33 in der Makula 444 eingebettet sind 444334, paarig 2. Und nun noch sogleich die Ohrsteine 55 auf die Hautstruktur 33 legen: #679813226185533. Stimmt fast! Hier wird zugleich die Zeilen-Nummer 553 adressiert, um die Steine für den Sacculus bereits herzustellen.
Kleines Vorhofsäckchen (Sacculus): Im Zeilensprung Zeile 553 zur Anlieferung der vorgefertigten Ohrsteinchen: #6601441274170550: es ist der Start des Sensororgan 66 zur Raumorietierung 44. Nach einigen Zeilen folgt die Struktur des Sacculus: #655403415550505: Die Evolution muss nehmen, was etwa sinnvoll als Code nutzbar ist: Sensor 6 als Struktur zur Lageorientierung 4 als Hautorgan 3 mit 4_03_4, bestehend aus dem Raumsensor 4 als Fasergruppe 1555 Haar 1 in einer Fasermenge 555, in der lotrechten Strukturmessung 0505. Die Härchen 333 werden dann angezeigt: #639333979792699. Und auf die Galertartige Sensorblase 699 hingeweisen. Die dann als herausgehobene Eleent 999 erscheint: #635992512999358.
Bogengänge (Canalis semicircularis): Der Start jeder der 3 Bogengänge ist an der Schlauchform 000 erkennbar. 1. Bogengang Zeile 686: #532157000672932. 2. Bogengang: # 421547000533062: Gleichgewichtsmessfeld: #418646447777100. Sensor 6_4_6 für die Drehbeschleunigung 100. Zeile 878: Das für den 1. Bogengang benötigte Sensorelement 1111 wird jetzt erst nachgeliefert: #415785538111197. 3. Bogengang: #405171700845318: Der Code 7008 hat ein Null weniger. Weil der horizontale und vertikale Gang eine teilweise gemeinsame Führung besitzen, muss es der frontale Gang sein (der hintere). Wegen der Unterschiedlichkeit muss die Evolution auch auf eine andere Codeart für das Gleichgewichtselement warten: So kommt unauffällig nach den Haaren #333 und der blasenartigen 000 Umhüllungsmasse #20003 dann als Besonderheit die aufgeweitete 000 Hauthülle des Bogenganges (Ampulla ossea lateralis): #3933833000664130. Weil die auffällige Ausformung bei den anderen Gängen fehlt, wird diese 3fach mit #3 adressiert sein. [Ergänzung Utriculus/Sacculus_dat06.01.2021]
In den sich anschließenden Code-Zeilen beginnt die Ausbildung der Brust. Hier interessiert der Geschlechter-Unterschied. Dazu hier weiter im BiXnom-Register „Gravitation“ zum Thema „Der Nippel“. Zur Artenvielfalt siehe BiXnom-Register "Prehistoric Slime / Urschleim" mit dem BiXnom-Baum-Code.
Auslese-Beispiel: Epi.ANN: Nase und Zunge
Die Nase gliedert sich in die äußere Nase (Nasus externus) und die innere Nase (Nasus internus). Die äußere Nase besteht aus dem festen knöchernen Nasenteil (Nasenpyramide), der auf jeder Seite nach oben den Nasenfortsatz des Stirnbeins umfasst. Als Überzug ist ein Schleimhautmantel vorhanden. Weiterhin seitlich den Stirnfortsatz des Oberkiefers und in der Mitte das Nasenbein (Os nasale). Der knorplige Nasenteil ist beweglich und besteht auf beiden Seiten aus dem Dreieckknorpel (Cartilago nasi lateralis) und dem Nasenspitzenknorpel (Cartilago alaris major). Die beiden Nasenlöcher (Nares) werden von den Nasenflügeln (Crus laterale) und dem mittleren Nasensteg (Crus mediale) begrenzt. Die knöchernen Anteile formen die Nasenwurzel, zusammengesetzt aus dem Os nasale, der Maxilla und dem Os frontale. Der darunter liegende knorplige Anteil besteht aus folgenden Knorpelelementen: Nasenspitze aus Cartilagines alares, Nasenrücken aus Cartilago alaris lateralis und Trennelement aus Cartilago septi nasi. Der zwischen beiden Nasenlöchern befindliche Nasensteg (Columell) ist an der Nasenscheidewand befestigt. Der Nasensteg wird durch Flügelknorpel stabilisiert, die die Nasenlöcher offen halten. Durch die Trennung der Nasenhöhle ist ein rhythmisches An– und Abschwellen möglich, d.h. eine Seite kann pausieren. Die innere Nase ist durch die Nasenlöcher zugänglich. Die Nasenhöhle (Cavum nasi), wird mittig durch die Nasenscheidewand (Septum nasi) getrennt. Auf jeder Seite gliedert sich die Nasenhöhle in den Nasenvorhof (Vestibulum nasi) und mittels bogenförmiger Schleimhautfalte (Limes nasi) abgegrenzter Nasenhaupthöhle.
Die Nasenmuscheln sind mit Nasenschleimhaut überzogene Knochenlamellen, die von der seitlichen Nasenwand in die Nasenhaupthöhle ragen. Sie zählen zur inneren Nase: #12124650198548400: Nasenmuscheln werden als paarige 2 Sensoren 6 zum Hirnnerv 1 als Struktur avisiert. 1+2= 3 auch 3 Sensoren zu lesen. Unterschieden werden 3 paarige Nasenmuscheln, zwischen denen sich die Nasengänge (Meati nasi) befinden. Für die obere und mittlere Nasenmuschel ist das Siebbein die knöcherne Grundlage:
Obere Nasenmuschel (Concha nasalis superior):#9093487648911300/4Hz: Sensor 11, herausgehoben 9, Nerv 1, paarig indirekt durch Doppeleins. keine Struktur 5, weil Bestandteil des Siebbeins. Die Nasenmuschel nimmt einen Teil der Riechzellen auf.
Mittlere Nasenmuschel (Concha nasalis media): #7274790119129040/5Hz: Sensor 11, herausgehoben 9, Nerv 1, paarig 2; keine Struktur 5, weil Bestandteil des Siebbeins. Die Nasenmuschel ist Teil des Eingangs zu den Nasennebenhöhlen.
Die untere Nasenmuschel (Concha nasalis inferior) wird von einem eigenen Knochen gebildet, der ebenfalls als Concha nasalis inferior bezeichnet wird. Die Nasenmuscheln nehmen nach oben in der Größe zu. Die untere Nasenmuschel ist besonders für die Luftdurchlässigkeit der Nase wichtig. :#30311625496371/12Hz
Riechgang: Zwischen dem Nasendach, den 3 Nasenmuschel und dem Nasenboden bilden sich 4 Räume, wovon der obere den Riechgang bildet und zum oberen Nasengang (Meatus nasi superior) gehört. Hier münden die hinteren Siebbeinzellen und - im Bereich des Recessus sphenoethmoidalis - die Keilbeinhöhlen (Sinus sphenoidales). Die Riechschleimhaut (Regio olfactoria) befindet sich am Nasendach über der oberen Nasenmuschel und in den obersten Abschnitten der Nasenscheidewand. Rezeptoren nehmen dort Duftmolekülen aus der eingeatmeten Luft auf. Die olifaktorischen Reize werden über Riechnerven zum Riechzentrum im Gehirn geleitet.
Sinusgang: Der mittlere Nasengang wird durch die Vertiefung der mittleren Nasenmuschel gebildet. In ihn münden über den Recessus frontalis die Nasennebenhöhlen, genauer gesagt der Sinus frontalis, der Sinus maxillaris und die vorderen und mittleren Siebbeinzellen. Der Gang wird deshalb auch "Sinusgang" genannt.
Atemgang: Der untere Nasengang führt zwischen der unteren Nasenmuschel und dem Nasenboden entlang und ist die kürzeste Verbindung zur hinteren Öffnung der Nasenhöhle. Der Tränengang (Ductus nasolacrimalis) mündet über die Hasner-Klappe in den Gang.
Riechkolben: Der Riechkolben ist Teil der Riechbahn. Die Riechnerven beginnen in der Nasenschleimhaut des Nasendaches und durchziehen das Siebbein (Os ethmoidale), das zum Hirnschädel gezählt wird, und sind mit dem Riechkolben (Bulbus olfactorius) verbunden. Es ist ein Anschwellung an der vorderen Basis des Gehirns und ein Teil des Riechhirns (Rhinencephalons) bzw. der Riechbahn. Im BiXnom-Code wird sich in der Nähe des Riechkolbens auch ein Hinweis auf das Neuroepithel befinden. Im Anschluss an die Nasenmuscheln ist 6fach die 33 für die Nasenschleimhaut vermerkt (33 hier kein Testosteron). Der Riechkolben führt in die Schleimhaut sensorische Fasern ein, die als eine 1-2 cm² große Region das Neuroepithel bilden. Das olfaktorische Neuroepithel besteht aus 3 zellulären Komponenten: den olfaktorischen Rezeptorneuronen (olfactory receptor neurons), den Stützzellen (supporting cells) und den Basalzellen. Letztere sind nochmals in die zwei Subtypen untergliedert: Neuroepithel: #559599239933003/65Hz als Struktur 55, deren Fasern gangartig 00 in hausschichten angelegt sind und durch das Siebbein mit 99 für herausgehoben durchzuleiten sind. Es folgen dann die 3 Zelltypen des Epithels, weil diese die 33 enthalten müssen: #466332699944169/78Hz [mit 999 als langfaserige ORN-Zellen]; #433023221376728/84Hz [23/22 als 2 Varianten GBC/HBC]; #413340347677786/88Hz [Stützzellen, 0-oberflächennah]. Der Riechnerv 1 ist adressiert/ es sind Fasern, so dass keine 5 als Struktur enthalten ist / die 6 weist auf die Steuerung durch das Nervensystem hin. Damit ist die folgende Struktur der Riechkolben: #404155006618280/90Hz. Blasenartige 00 Struktur 55 des Nervensystems 66 am 1. Hirnnerv. Die Ausstattung 8_2_8 zeigt mit der 2 eine Besonderheit entweder die folgende Stufe für das 2. Neuron oder die gemeinsame Verarbeitung mit den noch folgenden Geschmackswahrnehmungen.
Riechbahn (olfactory pathway): Als Riechbahn werden speziell-viszerosensible Fasern bezeichnet der Nasenschleimhau bis zu den olfaktorischen Cortexarealen des Großhirn. Es sind 2 Abschnitte vorhanden: Erstes Neuron: Von den Riechzellen des Nasenschleimhaut zum Riechkolben bildet sich der 1. Hirnnerv (Nervus olfactorius). Zweites Neuron: es befindet sich im Riechkolben, wo eine erste Reizverarbeitung durch Mitralzellen und Büschelzellen erfolgt. Die Reize werden über den Tractus olfactorius zur Riechrinde (olfaktorischer Cortex) mit der Area piriformis und dem medialen Teil des Corpus amygdaloideum geleitet. Der Riechstrang hat folgenden Verlauf: Riechstrang: Dem Riechkolben folgt der Riechstrang (Tractus olfactorius ), der die Verbindung zum Trigonum olfactorium als eine dreieckige Verbreiterung des Riechstranges ausgebildet ist. Es ist die Teilungsstellen in die Stria olfactoria medialis und die Stria olfactoria lateralis, die allein als die Fortsetzung des Riechstranges gilt, will die Stria olfactoria medialis vermutlich keine Nervenfasern enthält. Es wird eine Verbindung in die primäre Riechrinde hergestellt, in der der Geruch ins Bewusstsein gelangt.
Primäre Riechrinde: Die Stria olfactoria lateralis projiziert auf folgende Felder: Area pirifomis, Area entorhinalis und Amygdala. Die Stria olfactoria medialis projiziert auf Tuberculum olfactorium und Area septalis
Sekundäre Riechrinde: Der orbitofrontale Cortex überlappt die Geschmacks -und Geruchsinformationen.
Als bekannte Strukturen kann man nur den Mandelkern als vorläufig fixieren und dann die Hohlräume Nasennebenhöhle, Stirnbeinhöhle, Keilbeinhöhe suchen, um die Inhaltsbereiche des Programmcode zunächst als Übersicht auszulesen. Mandelkern (Amygdala): #410541203111119/886Hz: größte Signallänge 11111, herausgehoben 9, aus Zellen 3, Hirnnerv 1, paarig 2.
Das andere Ende des Suchweges bildet die Keilbeinhöhle: Die Keilbeinhöhle (Sinus sphenoidalis) gehört zu den Nasennebenhöhlen, aufgrund der Nähe zum Riechkolben zu fixieren: #363739505956452/100Hz: Ein Hohlraum als solcher ist nicht direkt vorhanden, aber die 505 ist ein Loch in der Knochenstruktur der Keilbeinhöhle (Sinus sphenoidales) als die Öffnung (Apertura sinus sphenoidales) zum hinteren Nasenraum, 52 ist die paarige Ausbildung der Keilbeinhöhle.
Erwartungsgemäß wird die Mitte zwischen beiden Höhle ausgelesen als eine Struktur 5 zwischen 2 getrennten Schleimhautarealen. Es ist unverkennbar die Nasenscheidewand (Septum nasi): #353145151414031/103Hz. Sie besteht aus der häutigen Pars membranacea #353, dem Nasenscheidewandknorpel (Cartilago septi nasi) 414, einem oberen knöchernen Anteil des Siebbeins (Lamina perpendicularis ossis ethmoidalis) #145 und einem unteren knöchernen Anteil, dem Pflugscharbein (Vomer) #514. Als höhergesetzte Struktur kommt nunmehr die Stirnhöhle (Sinus maxillaris) in Betracht: #279799619966501/130Hz: paarig 2, über 9 der Nase als Form 7 gelegen 7_9_7. Struktur 5 als Höhle 0 am Riechnerv 1. Es existiert weiterhin noch das ethmoide Labyrinth des Ethmoidknochens, das aus abgeschlossenen Luftkammern besteht und nur teilweise Bestandteil der Nasennebenhöhlen ist. Die gangartige Periode 222 in der mittleren Position würde für diese Funktion sprechen: #316295222570828/115Hz. Nachfolgend müssten die tiefer gelegenen Kiefernhöhlen folgen. Für einen Ausgang und die Höhle Ansicht werden 2 Nullen benötigt: #265503289019308/137Hz: paarige 2 Struktur 55 im sensorischen System 6; eine Höhle mit der 1 als Trennwand 9019, spiegelbildlich 03_30 ist die Nasen-Kiefer-Öffnung (Apertura nasomaxillaris) zur Nasenhöhle angezeigt. Die Öffnung mündet in den mittleren Nasengang. Weil es nur 3 Gänge gibt ist der Bezug #2 als der mittlere adressiert.
Als sekundärer Reiz wird der Geschmackssinn mit dem Geruchssinn überlagert. Es gibt 5 Geschmackswahrnehmungen: sauer, bitter, salzig, süß, umami (herzhaft-intensiv, fleischig). Die Rezeptorzellen für verschiedene Geschmacksknospen (Caliculi gustatorii) der Zunge in den Geschmackspapillen (Papillae gustatoriae), und auch in den Schleimhäuten von Mundhöhle, Rachen und Schlund angeordnet. Das muss dann die Zunge sein! #219120184311116/166Hz. Mit 1111 als langer Sensor und die Struktur 5 fehlt, weil die Zunge beweglich ist. Die samtartige Oberfläche geht von den mechanischen Papillen aus, die nach der Gestalt unterschieden werden in fadenförmige 11 und linsenförmige 0_0 Papillen. Die Zungenoberfläche wird adressiert: #217808087399073/167Hz und dann werden die beiden Papillen-Typen spezifiziert: #216511610688364 und #215230476897309. Es zählt die 1 für den Riechnerv 1 und dafür sind exakt nur noch diese 5 Programmzeilen verfügbar: #215230476897309/169Hz: umami Zungenmitte; #213964415268501/170Hz salzig Vorn seitlich 1_1; #212713161378042/171Hz: süße Spitze 1_1_1; #211476456951426/172Hz: bitter Mitte hinten 1 an 5; #210254049685810/173Hz sauer Außenrand hinten 8_5_8. Von den Geschmacksknospen führen afferente Fasern, die sich auf 2 Hirnnerven verteilen, und zwar auf den Gesichtsnerv (Facialis) und Zungen-Schlund-Nerv (Glossopharyngeus). Dabei könnte der Verzweigungspunkt aus folgender Anweisung resultieren: #155444233314723/234Hz. Die Verzweigung 2 erfolgt nicht in der Art von 333 und 444, sondern in der Programmzeile davor wurde offensichtlich der Gesichtsnerv auf die parallele Episode KRI ausgeleitet, so dass jetzt der Zunge-Schlund-Nerv (Glossopharyngeus) weiter geführt wird: Ausleitung bei Epi.ANN: #156111375946975/233Hz. In der Zeile 234 muss daher vorn #155444 als Achtungszeichen gelesen werden 5+4= 9 = 9. Hirnnerv IX. Der Nervus glossopharyngeus innerviert Zunge und Rachen und verfügt über sensible, sensorische, motorische und parasympathische Anteile. Die Ausleitung erfolgte nicht über dieselbe Programmzeilenummer vis-a-vis, sondern in unvorstellbarer Weise über die Programmzeileninhalte 156, der vorn wie eine Zeilennummer behandelt wird: EPI.KRI.Zeile 234: #156008419855398/234Hz. _dat02.01.2021
Auslese-Beispiel: Epi.VIR: Der virale Eingang
Als Eingang wirkt eine Erkrankung, wobei während der Genesung Strukturen frisch reproduziert werden, um schließlich die Heilung dem Gehirn zu vermelden: Wir sind nunmehr ein RNA-Stück der Form 777. Benötigt wird eine Virenhülle mit Geburtskanal als Spikes. Die 77 ist nun hinten rechts am Ausgang 00 in einer Virenhülle als Struktur verpackt: #656099003177. Wir stehen nun als 7 unten bereit und oben soll der Spikes mit 1111 zum Kontakt wedeln. Frisch geboren (zuvor war die Zeit noch negativ), steht die 7 jetzt vorn #76600466806. Es ist keine 5 für eine Struktur vorhanden, wir bewegen uns.
In der Geburtsphase ist die Zeit noch negativ. Die Gegenspieler sind die Leukozyten, die aber ebenfalls erst auf das Spielfeld gebracht werden müssen. Die Milz bringt Blut und Lymphe in einem Organ zusammen. Weil die Milz bisher noch nicht passiert wurde, wird sie ab EPI.VIR.Z.467 (Zeit -0,28Tage) bis Z.502 erwartet: #132345555071577. Die 1 für 1 Organkapsel als eine Hautkonstruktion die 2 Teile enthält, und räumlich 4 durch sogenannte Trabekel in Segmente unterteilt wird, die zur netzartigen Stabilität 5555 beitragen. Es ist eine runde 0 Form 7 als Struktur 5. Die 2 Bereiche sind die rote und die weiße Pulpa. Die Milz ist an 4 Haltebändern befestigt: #131781181702402. Mit 1 werden die Bänder adressiert, die die Milz-Hautkapsel tragen 1_3_1. Das sind 2 Bänder. Die Symmetrie meint dann 4 Bänder: 781_1_817. Die Gefäße zur Blutversorgung der roten Pulpa werden gebündelt über einen Gefäßstiel (Milzhilus) in die Milz geführt: #132062765423988: Die 1 ist hier der Stiel mit der 2 als Arterie und Vene. Arterie (Arteria splenica): #131500796209418: schlauchartige Struktur 500 als Hautgebilde 1_3_1. Die Folgezeile zeigt die Vene (Vena splenica): #131221601313007. Entsprechend dem Blutfluss rutscht die 131 nach hinten. Die Arterie verzweigt sich umgeben von der weißen Pulpa zur roten Pulpa, die über die Vene entsorgt wird. Die Stielgefäße verzweigen (gemäß Bildern) auf 4 Stränge: #130943589445819. Milzfollikel:#129030008806736. Weitere kleinere Bläschenstrukturen 0 sind vorhanden. Der Lymphabfluss erfolgt über Milzlymphknoten:#123118275335511. Der Knoten als 11, 33 hier als Lymphe, auch als Wirkstoff bekannt. Die Milz hat nunmehr die B- und T-Leukozyten für die Immunantwort bereitgestellt.
Die Zeitmarke TIME.Epi.VIR ist genau auf Geschlecht und Alter unserer Zielperson ausgerichtet, die unbedingt gesund bleiben muss. Nach 6,7 Tagen ist die Zielperson notgedrungen leider infiziert worden, sorry: #69655555. Oben 9 den Kopf erregt 6, das ist Fieber. Nach 14-tägiger Krankheit sind wir als Virus mit #350092750755786 strukturell in Bestform [die 7_55_7 hat als Zange die Struktur 55 im Griff] und warten nun auf die Begegnung mit den Leukozyten, die uns einen weiteren Einblick in den Körper gewähren sollen. Die Leukozyten antworten ebenfalls mit einem Klammergriff 7_55_7, der den Virus jetzt als 55 nach 14,03 Tagen gepackt hat. Die Leukozyten als Struktur 35 fressen den Virus nach 14,04 Tagen auf: #35005309367029: Der beseitigte Virus ist die Hautstruktur 53 mittig in der Blase 00_0_0. Die Leukozyten melden ihren Sieg nach 14,66Tagen: #349439555709993. Die Überreste des Virus werden als 9993 rechts aus dem Ziffernbild geschoben. Zugleich wird links der weitere Prozessverlauf als Lymphsystem mit 349 adressiert. Insgesamt zieht sich der Prozess über ca. 25000 Programmzeilen. Die Lymphe landet nach 14,47 Tagen erwartungsgemäß in der großen Lymphzisterne als Sammelraum im Bauch: #340000958402610. Das Lymphsystem ist wie adressiert unter 34_9 zu erkennen. Nach 15,3 Tagen liegt die Thymusdrüse vor: #322272261020056: Der Thymus ist ein zweilappiges 2 Organ. Die Evolution ist in der Psychologie nicht minder geschult als der Mensch. Wenn wir das Ende des Lymphsystems als den Übergabepunkt in den Blutkreislauf als Venendreieck bezeichnen, kann die Evolution das nach 16,21 Tagen ebenfalls mit der Ziffer 3: #304955712332493: Beidseitige 3 meint die beidseitigen Venendreiecke als Struktur 55. Die 233 bedeuten paarige Ausbildung der Venendreiecke. Es verbleibt die 1 für links und die 2 für rechts. Die 9 ist die Einleitung in die Vene von oberhalb. Nach 17,118 Tagen kann das Herz in vielen Programmzeilen bewundert werden: #289500090020265. Nachfolgend geht es bereits zur Lunge. Über den Blutkreislauf kann weiteres später ergänzt werden. Nach vielen Programmzeilen stabilisiert sich die kranke Lunge und zeigt nach 18,5 Tagen beide Flügel als Struktur an: #268508880955888.
Lunge? Das ist zunächst nur ein frommer Wunsch. Für die Lungenarterie (02arm) muss es im Herz zunächst die Pulmonalklappe geben: ab EPI.VIR.Z.22920: #269656955577777: 555 drei halbmondförmige Elemente. Nach der Lunge geht es mit 2 Öffnungen für die beiden Lungenvenen (02reich) durch die Herzwand in den linken Vorhof zurück: EPI.VIR.Z.23089: #267683200738128: 1=links. Linker Vorhof: #267660015670289. Mitralklappe zur linken Herzkammer: #267555732547300. Linke Herzkammer: #267509410571444: Struktur 5, Form 7, links 1, räumlich weit 44_4 pulsierend. Aortenklappe: #267486255597795. Beginn der Aorta: #2674631046322760: 6_74_6 mit 7+4= 11 folgt 6_11_6_310. Im Prinzip ist gemeint, dass oben 1 Öffnungsloch 0 codiert ist. Der senkrechte Verlauf der Aorta (Pars ascendens aortae) wird sofort als Bogen (Arcus aorta) mit Richtungswechsel nach unten (Aorta thoracica) verändert, wobei 3 Arterien im Bogen nach oben abgehen sollen: #267439957673849. Der Bogen ist 3x die 7 und mittig hoch mit 99. Die 3 Abhänge werden angekündigt mit der 8 als Extras. Drei arterielle Abgänge nach oben 9 als Gefäß 3 Extras 8: #267428385696969: 69_69_69. Die unterschiedliche Lage wird codiert: #267416814721471: Die 1 beschreibt, dass mit der Symmetrie 41_68_14 die Halsschlagader 1 nach oben zum Hals zu führen ist. Die 2 am linken Ziffernrand ist "rechts" und die 1 am rechten Ziffernrand ist "links", so dass die Reihenfolge bestimmt ist. 41 führt zur rechten Seite während 14_72_14 links orientiert bleibt. Im Ablauf muss dann die Halsschlagader ausgewählt werden: #267382107801974.
Die 3 Äste des Aortenbogens (Arcus aortae): 1.) Truncus brachiocephalicus (Arm-Kopf-Gefäßstamm): Versorgung von rechten Arm und der rechten Kopf-, Hals- und Gehirnhälfte mit arteriellem Blut. Er bildet den gemeinsamen Gefäßstamm der rechten Arteria subclavia und der rechten Arteria carotis communis. 2.) Arteria carotis communis sinistra (links): Die Arteria carotis communis (Halsschlagader) liegt beidseitig. Vom Aortenbogen geht der linke Zweig ab, der rechte vom Truncus brachiocephalicus. Kopf und Hals werden mit arteriellen Blut versorgt. An der sogenannten Karotisgabel (Bifurcatio carotidis) teilt sich das Gefäß in: 2.1) Arteria carotis interna (ACI): versorgt den vorderen Teil des Gehirns, das Auge, seine Anhänge und bildet kleinere Äste zu Stirn/Nase und 2.2) Arteria carotis externa (ACE): Sie versorgt den größten Teil der Kopf- und Halsweichteile, sowie Teile des knöchernen Schädels und der Dura mater (äußerste Hirnhaut) mit arteriellem Blut. 3.) Arteria subclavia sinistra (links): Die Arteria subclavia ist eine paarig vorhandene stammnahe Arterie, die arterielles Blut zu Kopf, Hals, Arm und Schulter führt. Von den 4 Ästen die die Arterie bildet versorgt die Arteria vertebralis das Gehirn (Hirnstamm, Kleinhirn und okzipitale Abschnitte des Endhirns).
Die paarige Halsschlagader (Arteria carotis communis) wird rechts aus einem Abzweig des 1. Astes vom Aortabogen und links direkt vom 2. Ast gebildet und liegt beidseitig am Hals an: #267393675774104: Gefäß 3 liegt beidseitig am Hals 9: #6739367. Weil die Zeit nicht rückwärts läuft, muss in der Folgezeile der zweite arterielle Anschluss des Gehirns folgen. # 267382107801974. Die Arteria vertebralis („Wirbelarterie“) ist ein Ast der Schlüsselbeinarterie (Arteria subclavia). Sie zieht durch ein Loch im Seitenfortsatz der Halswirbel (Foramen transversarium) zum Schädel hoch: #267370540830708: das Gefäß 3 soll durch eine Form 7 geführt werden, und zwar durch die Halswirbel als Extras 8, die Löcher für die Durchleitung besitzen 070. Sie zieht durch das Foramen magnum und vereinigt sich mit der A. vertebralis der Gegenseite zur A. basilaris. Dieser Übergabepunkt verbleibt dann als: #267358974860174. Angezeigt wird eine Formzusammenführung auf 01 statt 11.
Die Verzweigung der Hauptschlagader (Arteria carotis communis) wird mit Verweis auf 8 avisiert: #267347409890243. Die Verzweigung (interna/externa) wird als Struktur 5 durch Zahlendreher 58_85 symbolisiert: #267335845920785. Der interne Zweig führt zum Gehirn. Die Ziffer 8 als Hauptschlagader weiter aktiviert, die jetzt durch den Knochenkanal Canalis caroticus mit #282 geführt wird (2 als Knochen): #267324282951671. Die Arterie verzweigt sich in die in die Arteria carotis externa (30% der Strömung) und die Arteria carotis interna (70% der Strömung): #267312720982771. Verhältnis 2/7= 0,29. In der letzten Zeile vor der Gehirnversorgung muss noch die Abgabe der A. ophthalmica codiert werden. Dieser Abzweig versorgt das Auge und Teile der Nasennebenhöhlen: #267301160013954. Anschluss an die sechseckige Circulus arteriosus cerebri (Willisi): 3_011_60 und mit 01_3_9_5_4 eine abgehende räumliche 4 Gefäßstruktur 3_5.
Aufgrund der sechseckigen Form passt hierzu die 6 allgemein für Steuerungsfunktionen: Epi.Vir.Z.23123: #2672896000450910. Für die Blutversorgung des Großhirns stehen 8 Zeilen #2666... zur Verfügung, die in direkter Reihenfolge auch verwendet werden:
Vordere Gehirnschlagader: Arteria cerebri anterior: vordere/oberer Teil des Gehirns: #266689856390189: 8_9_8= oben. Mittlere Gehirnschlagader: Arteria cerebri media: laterales (seitliches) Gehirn: #266678349233804: 4_9_2= seitlich. Hintere Gehirnschlagader: Arteria cerebri posterior: hinterer/unterer Teil des Gehirns: #266666843070399: Arteria cerebri anterior (aus der Arteria carotis interna): #266655337899846. Arteria communicans anterior (aus der Arteria cerebri anterior) : #266643833722017: Arteria cerebri media (aus der Arteria carotis interna) :#266632330536783: Arteria cerebri posterior (aus der Arteria basilaris):#266620828344016: Arteria communicans posterior (aus der Arteria cerebri anterior): #2666093271435870.
Die Venen können mit #665 wegen der 5 als Kanalstruktur 8 Elemente belegen. Der Venenhohlraum an 9. Stelle ist wegen der Nullen passend. Hirnvenen:1.) die oberflächlichen Venen (Vv. cerebri superficiales. 2.) tiefen Venen (Vv. cerebri profundae) 3.) Kleinhirnvenen (Vv. cerebelli) 4.) Venen des Hirnstamms (Vv. trunci encephali). Hirnsinus: Das Blut der Venen fließt aus allen Hirnbereichen 5.) in die Sinus durae matris (Sinus sagittalis superior, - rectus, - transversus, - sigmoideus und - cavernosus) ein: #266551836022023: Es wird zwischen einer oberen und unteren Gruppe 22 unterschieden. 6.) Der Hauptabfluss aus dem Schädel erfolgt mit der Vv. jugulares internae: #266540340772927. Es gibt noch 5 ergänzende Abflüsse. 7.) Der Sinus sagittalis superior entleert die oberflächlichen Venen des Gehirns. Das Drainagegebiet umfasst den oberen Teil der medialen und lateralen Hirnrinde des Frontal-, Parietal- und Okzipitallappens. Über die Vena anastomotica superior (Trolard-Vene) steht der Sinus sagittalis superior mit der Vena media superficialis cerebri in Verbindung. Der venöse Abfluss erfolgt über den Confluens sinuum via Sinus transversus und Sinus sigmoideus in die Vena jugularis interna: #266528846515272: Verbindung 2_7_2. 8.) Die Vv. superficiales cerebri leiten das Blut aus den obersten Schichten des Gehirns und sind über Brückenvenen mit dem über die Brückenvenen mit dem Sinus durae matris verbunden: #266517353248928: Brücke 3_5_3. Es sind 4 Unterteilungen vorhanden. Ein Kanal verläuft weiter nach dorsal und mündet 9.) in den Confluens sinuum, einem venösem Hohlraum 5_0_5 am Polus occipitalis ein: #266505860973767: Es ist ein venöser Zusammenfluss von 3 Hirnleitern, paarig 2.
Wird nun noch der Riechkolben (Bulbus olfactorius) mit Blut versorgt, kann er die Gesundmeldung des Körpers an das Gehirn weiterleiten. Es ist eine Ausziehung der Großhirnrinde, die wegen zu geringer Zellzahl aber selbst keine Informationen verarbeiten kann, sie werden aus der Nasenhaupthöhle kommend weitergeleitet. Der Riechkolben ist aus 6 Schichten aufgebaut: Nach 18,7 Tagen: #265841000552395. Die 1000 können die ca. 1000 genetisch definierten Rezeptortypen darstellen. Mehr Geruchswahrnehmungen entstehen durch Kombinationen. Der Nebenriechkolben (Bulbus olfactorius accessorius) 2 ist beim Menschen nur rudimentär vorhanden.
Nach 18,75 Tagen wird auf den Signalweg zum Gehirn verschaltet: #265111200696720. Und dann? Jetzt passt etwas Unglaubliches!!! Nie hat es hinter dem Komma eine glatte Tageszahl Tag-Komma-Null gegeben. Das geschieht nun exakt nach 19,0 Tagen! Was ist geschehen? Mathematik vorweg: Den Goldenen Schnitt kann man exponieren, ohne dass sich dabei die Nachkommastellen ,6180 ändern. Setzen wir nun also den großen Goldenen Schnitt mit PHI=1,6180...ins Quadrat als Quadratkopf namens Gehirn, dann sind das 2,6180... Und nun staunen: #261809523524490. Diese Gesundmeldung hat das Gehirn verstanden und ist als Fläche allen Gehirnarealen bekannt. Uns jetzt auch!
Der Körper-Rundgang ist weiterhin noch aktiv, weil nach der Genesung noch die Lymphknoten abschwellen müssen. Um den geeigneten Ziffernbereich für Knoten #111 an Kopf und Hals zu erreichen, sind nach 18,72 Tagen Programmzeilen ab EPI.VIR.Z.23272 geeignet, die Schilddrüse zu adressieren. Dazu passt sich die Evolution exakt 2 Programmzeilen an: #265578266665634. Die Schilddrüse besitzt 1 linken und 1 rechten Lappen 6666. 4x die 6 ist schon sehr breit, aber vorn steht die 2 als Doppel. Die 6 weist generell auf die Steuerungsfunktion hin. Die Schmetterlingsform wird durch einen Verbindungslappen in der Folgezeile erzeugt: #265566855233216: Beidseitig zentriert durch die 55, wird der Verbindungslappen 66 als besondere Ausstattung bzw. als Aufweitung erzeugt. Mit 33 wird hier nicht Testosteron erzeugt, sondern das Schilddrüsenhormon. Auf der Rückseite sitzen die 4 Nebenschilddrüsen als Viereck. Beide Reihen werden getrennt adressiert: #265555444781414: Es sind 2 Drüsenknoten als Doppelstruktur 55-55 und räumlich ausgedehnt 444, wobei der Abstand in der oberen Reihe größer sein soll, besondere Ausstattung 8, 14-14 meint, wegen der 4 als räumlich ausgedehnt, das vorn kein Platz wäre, daher auf der Rückseite als punktuell 1, als eine Form 7. Nun für die untere beide Drüsen: #265544035310103: Würfelkunde extrem! Wurde ein Organ durchlaufen, ist es nach der Virusinfektion wieder in Takt. Aber 353 bleibt schwer krank, gesund wäre 535. Über einige Programmzeilen muss der Patient daher als Placebo-Effekt genötigt werden, aufzustehen 999 /44 / statt 4 auf 2 Beinen stehen, um frische Luft zu atmen. Schließlich sehr nachdrücklich: #265429994496141. Egal wie, er muss stehen 141. Warum? #265373010813338: Für Vitamin D etwas Sonne als 333 tanken! Mit 535 ist die Hautstruktur nunmehr gesund und produziert nachfolgend Wirkstoffe: #265350224190393. Der virale Eingang mit einem Krankheitsverlauf an einem Goldenen-Mann-Modellkörper (er könnte in Dresden den „Goldenen Reiter“ als August der Starke abbilden) lässt die Organe mit weit über 20000 Programmzeilen gesunden. Daher ist es der Evolution möglich, auch Details wie Drüsen etc. auszuformen.
Bis zu den Lymphknoten sind aber 12 unbekannte Programmzeilen vorhanden. Die Evolution kann hier nur die Ohrmuscheln kreieren und muss das tun, weil mit #2653... eine Hautkonstruktion entstehen muss – es dürfen auch Segelohren sein. Sind das Ohren? #265338832346312: 33 ist eine Haut-Membran, 88 ist weit und eine besondere Ausstattung; 3_2_3 sind paarige Muschel, 4 räumlich, 6 _3_1 ist die Muschel 6, an der Haut 3 um 1 Gehörgang, paarig 2. Folgezeile: #265327441480323: Die Muschel kann hier als Raum 44 Breite/Höhe= 1:4 (passt, weil Schubverhältnis GS) geformt werden für 3_2_3 beide Muscheln. Der Ohrgang wird adressiert:#265316051592300. Ohrgang ist 5_1_5, soll wachsen 9, paarig 2 als lange Hautröhre 3_000. Nun kommt wie erwartet das Ohrläppchen mit der 11 und wegen der 9 herausgestellt an der Muschel: #265304662682119. Ohrläppchen 11 gehören zur Muschel 66_2682 ergibt 4 Form 8:2=4:1. Die Höhe ist die Länge 4, so dass nun die Muschel 6 in Relation 9 symmetrisch zum Ohrgang anzusetzen ist (Muschel hängt nicht): #265293274749652. Nach der Höhe wird die Breite 1 codiert 8_1_8: #2652818877947740. Der Muschelrand hat eine gedoppelte Form 77, davon oben beidseitig umgekrempelt 4_77_4 und unten nur nach innen 8_77_9. Mit 1_8_1 wird die Größe der Ohrlappen codiert: #265270501817359. Da Ohr verjüngt sich evtl. 70/50 nach unten. #265259116817281. Das Ohrläppchen 11 soll bezüglich der zwei 5_2_5 eine Symmetrie 81_7_2_81 ausbilden: #265259116817281. Es handelt sich mit der 81 und Tragus und Antitragus im oberen Bereich des Ohrläppchens. Es werden die Helix 77 und die Antihelix 44 codiert: #265247732794414. Es werden die oberen Gegenkrümmungen codiert: #265236349748633. Crus inferius inferius 6_7_6 und Crus helicis 22 : #265224967679811. Die Höhe 8:2=4 wird nochmals zur Breite 1 codiert : 5_86_5. Das ganze Ohr betreffend wird es sich um die Scapa (zwischen Helix und Anthelix) handeln: #265213586587824.
In EPI.VIR.Z.23305 wird die Bildung der Lymphknoten vorangekündigt: #265202206472544: Ausbildung paarig 2, gesteuert 6 als Immunantwort, Knotenstruktur 5, Form 7 rund mit der inneren Filterfunktion 0_22_0 als räunmliche Struktur 5_44. Mit der 2 werden nur die Kopf- und Halsknoten ausgebildet. Es wird die bekannte Riehenfolge übernommen. Für die Achsel-Lymphknoten reichen die Zeilen nicht aus, daher folgen sie später verschleppt über einen weiten Zeilenbereich. Als Knoten können sie jedoch wegen der 11 am Zahlenende identifiziert werden. In der Rückwärtssuche wird zuerst die Durchtrittsstelle zur Versorgung der Oberschenkel (Lacuna vasorum) mit Nerven und Blutgefäßen unter dem Leistenband gesucht. In der folgenden Programmzeile findet sich dann der letzte Lymphknoten.
Die 6 Lymphknoten des Kopfbereichs:
1.) Nodi lymphatici retroauriculares: #265190827333847: 2 kleine Knoten hinter der Ohrmuschel: Ziffernbild ist mit der 333 als Ohr-Schwingung korrekt.
2.) Nodi lymphatici parotidei: #265179449171607: Liegen im Bereich der Ohrspeicheldrüse, es wird zwischen einer oberflächlichen (Nodi lymphatici parotidei superficiales) und einer tiefen (Nodi lymphatici parotidei profundi) Gruppe unterschieden. 179 ist die tiefe Gruppe / 917 ist die Oberfläche/ 44 die Räumliche Ausweitung.
3.) Nodi lymphatici faciales: #265168071985698: Kleinere Knoten im Gesichtsbereich, die Lymphe weiter durchleiten 0, hier mit der 56 von 5 nach 6 symbolisiert.
4.) Nodi lymphatici occipitales: #265156695775994: 1-3 Knoten an der Rückseite des Kopfes zur Dränage der Kopfhaut. Im Ziffernbild ist die Kopfhaut als Membran 33 adressiert, so dass mit 2 Knoten die 66 entsteht. Die 5775 symboliert den Hinterkopf mit 5994 eine herausgehobene Struktur.
5.) Nodi lymphatici submandibulares: #265145320542370: 3-6 Lympkknoten im Gewebe hinter dem Unterkiefers (Dränage für Nase, Lippen, Mundschleimhaut, Zunge): Die Null steht für Mund/Nase und die Umkehrung 32>23 meint den Unterkiefer. Die Anzahl ist mit 4 bestimmt.
6.) Nodi lymphatici submentales: #265133946284700: 1-2 Knoten unter dem Kinn für Unterlippe und vordere Zunge: Die Zunge ist als räumliche 4 lange Form 7_00 bestimmt. Die Unterlippe mit 339. Knotenanzahl 2.
Die 2 Lymphknoten des Halsbereichs:
7.) Nodi lymphatici cervicales superficiales: #265122573002859: Sie dränieren die Haut und das Unterhautzellgewebe und leiten die Lymphe tiefer weiter. Sie liegen die oberhalb der oberflächlichen Halsfaszie, einer derben Hüllschicht aus Bindegewebe. Eine Faszie kann Muskelgrupen oder Körperabschnitte umgeben. Die 3 steht für die Haut und 00 für die Abflussröhre.
8.) Nodi lymphatici cervicales profundi: #265111200696720: Die Knoten liegen unterhalb der Halsfaszie und nehmen alle Lymphe oberhalb und des Kopfbereiches auf. Mit der 111 werden die 3 Knotengruppen adressiert: 8.1) vordere tiefe Halslymphknoten (Nodi lymphatici cervicales anteriores profundi) in der vorderen Halsgegend unterhalb des Zungenbeins, 8.2) seitliche tiefe Halslymphknoten (Nodi lymphatici cervicales laterales profundi) in der lateralen Halsgegend und im Bereich des Musculus sternocleidomastoideus. 8.3) retropharyngeale Halslymphknoten (Nodi lymphatici retropharyngeales).
Die 5 Lymphknoten des Brustraumes:
1.) Nodi lymphatici mediastinales: Das Mediastinum ist ein komplexer anatomischer Raum, der allmein adressiert wird mit: #265202206472544. Das Räumliche ist die 44. Die 0_22_0 ist das arteriellele Versorgungsgebiet. Nach der Lage werden 2 Lymphknotengruppen unterschieden:
a) Die vordere Lage mit den Nodi lymphatici mediastinales anteriores: #265179449171607. Mit dem zuvor adressierten Neetzwerk 44 werden die Knoten statt 11 als Bereich adressiert 1___1.
b) Die hintere Lage: Summen-Ziffernbild: #265111200696720. Mit der 2 kann ein 1 verdoppelt werden, so dass die Evolution sich den Goldenen Schnitt hier schön rechnet als 11.11 für 2 Untergruppen. Es können somit wie in der Radiologie auch 3 Gruppen erkannt werden. Andererseits 111 statt 11 auch 11_1 als hinten.
2.) Nodi lymphatici tracheobronchiales: Die Knoten liegen neben Luftröhre und Hauptbrochien: #265088459011050. Die 11 liegt nicht am rechten Zahlenrand, daher wird mit 0_11_0 die Luftröhre als Bereich avisiert. Unterschieden werden: a) Nodi lymphatici tracheobronchiales superiores: Sie liegen im Winkel zwischen der Trachea und den Hauptbronchien: #264793171751109. Beidseitig der Luftröhre als Ziffernbild 1_7_1. b)Nodi lymphatici tracheobronchiales inferiores: Sie liegen in der Tracheagabelung zwischen den beiden Hauptbronchien: Die Gabel wird durch die direkte Zeilenfolge ab EPI.VIR.Z.23345 realisiert:#264747801321167, Dopplung als Gabel; gefolgt von #264736461142921. Als Gabel liegt die 46 als Wert dazwischen 47_46_47.
3.) Nodi lymphatici interpectorales: Das Gebiet ist die vordere Brustwand incl. Brustdrüsen (Mamma): #264600454741101. Mit 4_600_4 wird die Brustdrüse und mit 44 das Räumliche (Busen) adressiert. Für die Brustwand noch 2x die 11 nicht am rechten Zahlenrand.
4.) Nodi lymphatici intercostales: Mehrere kleine Lymphknoten nahe der Rippenköpfe: #264080388901155. Keine 11 am Zahlenrand, sondern an der Struktur 55 der Rippenköpfe auszurichten.
5.) Nodi lymphatici parasternales: Die Knoten liegen neben dem Brustbein (Sternum) #263978875916911: mit 7_88_7 wird die symmetrische Lage des Brustbeins symbolisiert. 88 ist eine Weite für den Brustkorb, d.h. vorn.
Die 5 Lymphknoten des Bauchraumes:
1.) Nodi lymphatici coeliaci: Knoten im Bereich der Bauchaorta (Truncus coeliacus): #263764826811311: Es sind insgesamt 11 Knoten (10 x mit der 11 nicht am Zahlenende). Die Anzahl muss codiert sein, weil die Bauchaorta im Zeilenbereich dazwischen liegt: #263248037390009. 7 Knoten davor, 4 dahinter= 7_6_4 =7+4=11 und mit 2+6+3=11 wird die 6 als die Aorta codiert.
2.) Nodi lymphatici mesenterici superiores: #: Knoten um die Arteria mesenterica superior im Lendenwirbelbereich.
3.) Nodi lymphatici mesenterici inferiores: Knoten am Ursprung der Arteria mesenterica inferior (3. unpaarige Ast der Bauchaorta): #261267222769811. Mit der 222 als Artierienknoten.
4.) Nodi lymphatici iliaci externi: Es sind 8-10 Lymphknoten, die entlang von Gefäßästen lokalisiert sind (Arteria und Vena iliaca externa) in 3 Gruppen lokalisiert sind: Ab EPI.VIR.Z.23663 werden in direkter Folge 7 Knoten avisiert mit #11 für die 3. Gruppe ist als Besonderheit der 8.Knoten mit #111 vorgesehen: a) Nodi lymphatici iliaci externi laterales: Mehrere Lymphknoten entlang der Medialseite der Gefäße. b) Nodi lymphatici iliaci externi mediales: Mehrere Lymphknoten entlang der Lateralseite der Gefäße. c) Nodi lymphatici iliaci externi intermedii (Knoten liegen zwischen a/b): #261112692093056 und eine Vielzahl mit 11 nicht am Zahlenrand als „dazwischen“ zu verstehen.
5.) Nodi lymphatici iliaci interni: Knoten an den Gefäßästen der Arteria iliaca interna werden unterschieden in 3 Gruppen: a) Nodi lymphatici gluteales superiores: 2-3 Lymphknoten entlang der Medialseite der Arteria glutealis superior.: #259424841413811: mit 414 werden die folgenden 4 Knoten als 11, nicht am Zahlenrand vorangekündigt. b) Nodi lymphatici gluteales inferiores: Mehrere Lymphknoten entlang der Arteria glutealis inferior - bei der Frau zwischen Arteria uterina, Arteria rectalis media und Arteria vaginalis. Bei der hier gewählten Verteilung verbleibt 1 Programmzeile #259098575578211 für den Mann und dann als Alternative für die Frau: #259011709908752. Eine Zwischenposition des Gefäßbeeichs wird symbolisiert durch 0_99_0. c) Nodi lymphatici sacrales (laterales): 2-3 Lymphknoten lateral der Arteriae sacrales laterales:#259098575578211 incl 2 weiter mit 11, aber nicht am Ziffernrand.
Der Lendenlymphknoten (Nodi lymphatici lumbales): Die Lymphknotenstruktur 115 wird als räumliche Verteilung 44 angekündigt: EPI.VIR.Z.23955: #258006154115744. Das Gebiet umfasst eine Vielzahl von Organen. Nach der Lage werden 3 Gruppen unterschieden: Die Evolution unterscheidet, ob die 11 im Ziffernbilde rechts, mittig oder links liegt: Nodi lymphatici lumbales dextri (rechte Gruppe): #25799538411432; Nodi lymphatici lumbales intermedii (mittlere Gruppe) zwischen Aorta und Vena cava (Hohlvene): #257243711888897; Nodi lymphatici lumbales sinistri (linke Gruppe):#257211595232537: Mit 3 Untergruppen: seitlich 572, vor 523 und hinter 253 der Aorta. Arterie mit 2 adressiert.
Der Leistenlymphknoten (Nodi lymphatici inguinales): Unterschieden werden die oberflächlichen (Nodi lymphatici inguinales superficiales): #257115293362286: 115 links ist oberflächlich; und die tiefen (Nodi lymphatici inguinales profundi) Leistenlymphknoten: #256901547171113. Zu letzteren gehört auch der „Rosenmüller-Lymphknoten“ in auffälliger Größe in der „Lacuna vasorum (Durchtrittsstelle für Gefäße)“ 7_1_7: EPI.VIR.Z.24037: #257125990008846: Arterie, Vene, Nerv =000. _dat03.01.2021
Das Universum - Entstehung der Atome aus Episoden.BIX/XIB
Die Biologie kann den gewölbten Randbereich der BiXnom-Gravitationslinse durch die Fähigkeit nutzen, auf dem Wege der Evolution Strukturen auszuformen, die exakt dem Goldenen Schnitt entsprechen (Energieminimum). Die Teilchen können dagegen nur unflexibel den rechteckförmigen Bereich ausfüllen. Die Strukturgrundlage ist somit selbstverständlich erst recht der Goldene Schnitt. Der Goldene Schnitt zeigt im dynamischen Ablauf ein 3er Muster beim Zeitvorlauf (0/5; 0,333; 0,666). Die Reizleitung erfolgt im Goldenen Schnitt bevorzugt mit 11 etc. Die Bausteine der Atome sind eine 3er Gruppe und folgen deshalb bei der Strukturbildung dem Signal 111. Der Goldene Schnitt bietet als bevorzugte Strukturbildungsplätze das Signal 111.xx an, und zwar bis zur 5 Ebene des Periodensystems, darüber hinaus muss das Signal noch stärker wirken. Die Perioden 1 - 5 sind ohne Besonderheit zu erzeugen! Die lotrechten Gruppen 3 bis 12 mit jeweils 10 Sekundärplätzen liegen zwischen 1 und 2 sowie 13 bis 18. Zur Besetzung der Sekundärplätze werden die weniger markanten Signale verwendet. Insgesamt werden alle deutlichen Signale verwertet.
Die Atome entstehen aus der Episode BIX und der Episode XIB. Zunächst muss das Universum aus dem Nichts entstehen. Das geschieht hier mit der Bildung eines Klopses (wie bei der Niere) unter Verwendung der Steuerkugel. Damit hat Wasserstoff (H) seinen Platz im Goldenen Schnitt gefunden. Die Episode EPI.BIX hat in der gleichen Programmzeile ein Ergebnis, und dadurch bildet das nun entstandene Helium (He) mit dem Wasserstoff ausnahmsweise eine gemeinsame Periode. Es stehen 8 Primärplätze zur Verfügung. EPI.XIB erzeugt die 3. Periode. Es stehen 8 Primärplätze zur Verfügung. In der 4. [EPI.BIX] und 5. Periode [EPI.XIB] stehen je 8 Primärplätze und 10 Sekundärplätze zur Verfügung. Die Plätze bestehen aus einer Ziffernfolge, die keinen direkten Massebezug enthält. Entsprechend dem Zeitablauf ist die Reihenfolge der Plätze bekannt. Die Plätze können sich zeitlich überschneiden. Entweder Epi.BIX hat als führender Vorgang die Vorfahrt oder es muss im Zweifelsfall das stärkere Signal entscheiden. Eventuell sind in den Programmzeilen auch Sprungmarken enthalten, die den Vorrang steuern. Im Gegensatz zur Evolution steckt dann keine "Denkleistung" dahinter, sondern der Goldene Schnitt muss mit seiner Zahlenfolge eingreifen.
TIME.EPI.BIX = Radius BiXnom - Kugel abzüglich Radius Signalkugel; TIME.EPI.XIB = Radius BiXnom - Kugel abzüglich Radius Steuerkugel (= halbe Signalkugel).
Der Radius der BiXnom - Kugel ist die Differenz zwischen der Sehnenhöhe aus dem BiXnom-Bogen einer Sehne der Länge 1 in Höhe y= -0,25 und der Oberkante des Quadrats y=0. Die BiXnom-Kugel fällt der Gravitation folgend lotrecht in den CC-Point (CCP). Der Aufschlag auf die dortige Signalkugel bzw. die Steuerkugel erzeugt eine Folge von Signalen über die Umdrehungen im BiXnom-Schwingkreis.
Bei der 6. Periode, beginnend mit Caesium (Cs), hat sich die Länge (Zeit) und damit der Signalabstand bereits extrem verkürzt, so dass im Gewitter die vielen #111-Signale nicht mehr trennscharf sind. Daher wird jetzt das stärkere Signal #1111 für die Bestückung des Periodensystems der Elemente (PSE) verwendet. Es können ebenfalls wieder 8 Primärplätze und 10 Sekundärplätze bereit gestellt werden, allerdings verbunden mit einem Unfall: Der Vorlauf der #111-Perioden konnte ignoriert werden, aber für den Nachlauf, die den # 1111 mit #111 folgen, fehlt das Stopp-Signal: Die Gruppe der Lanthanoide, beginnend mit Cer (Ce) mit 14 Extra-Plätzen wird deshalb nun ins Periodensystem eingelesen. Die Warteschläge der #111 ist noch lang, aber es kommt das einzige Stopp-Signal, welches Teilchen verstehen können, es ist der signalisierte Goldene Schnitt: #1110615... mit phi = 0,618... In der 7. Periode kann ein Ergebnis gefunden werden, weil die Anzahl der Elemente bekannt ist. Zum BiXnom-System gehört das natürliche Gleichgewicht, zum PSE gehören aber auch künstliche Elemente. Dem Anschein nach folgt nach den ersten beiden markanten Elementen eine Wende. Die 10 Elemente für die 3.-12. Gruppe können gefunden werden, wenn nach der #1111 direkt eine Doppelziffer folgt. Bei den weiteren Elementen muss eine Auswahl getroffen werden. Im Gegensatz zu den Kristallen ist die 5 beim Atom keine Strukturvoraussetzung. Im Gegenteil wurde die Auswahl so getroffen, dass das Energienieveau sehr tief liegt. Die maximale Doppelziffer in der mittleren bis hinteren Ziffernfolge ist 44 < 55. Es fallen dann 20 Elemente an, von denen nochmals 6 für die Hauptgruppen 13-18 entnommen werden können, so dass die Gruppe der Aktinoide mit 14 Elementen besetzt ist.
Mit der Entstehung der Kristallsysteme incl. Bravais-Gitter als ein freier Fall der BiXnom-Kugel auf die Untergrenze wird am Ende Register „Surprise“ fortgesetzt._dat24.01.2021
Chris Acker: PANICKED AND PARALYZED